

Tri charakteristiky a funkcie v organizme
- 4743
- 606
- Adrián Fajnor
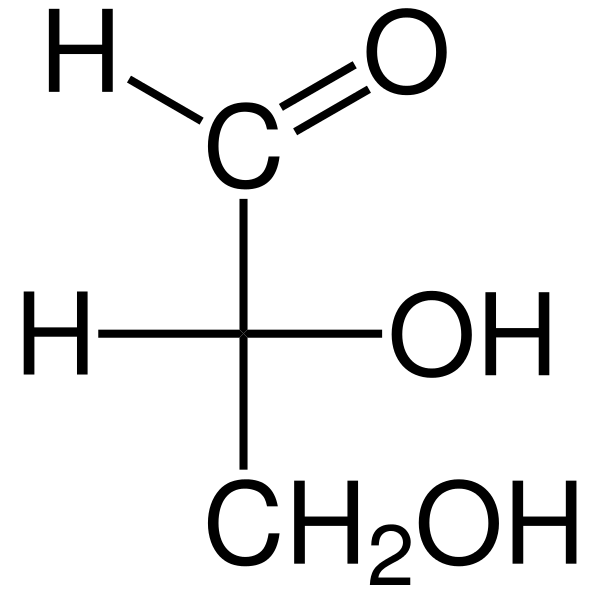
Ten Triosas Sú to tri -karbonové monosacharidy, ktorých empirický chemický vzorec je C3H6Ani6. Existujú dva triosy: glyceraldehyd. Triosy sú dôležité v metabolizme, pretože spájajú tri metabolické dráhy: glykolýza, glukoneogenéza a dráha fosfátu penózy.
Počas fotosyntézy je cyklus Calvin zdrojom triosov, ktoré slúžia pre biosyntézu fruktózy-6-fosfátu. Tento cukor fosforylovaným spôsobom sa premieňa enzymaticky katalyzovanými krokmi do rezervných alebo štrukturálnych polysacharidov.
Zdroj: Wesalius [verejná doména] Triosy sa zúčastňujú na biosyntéze lipidov, ktoré sú súčasťou bunkových membrán a adipocytov.
[TOC]
Charakteristika
Glyceraldehyd aldosa má atóm királneho uhlíka, a preto má dva enantiomeros, l-glyceraldehyd a d-glyceraldehyd. Obidve enantioméry d a l majú rôzne chemické a fyzikálne vlastnosti.
D-Glyceraldehyd prelomil rovinu polarizovaného svetla doprava (+) a má rotáciu [α] D, pri 25 ° C, od +8.7 °, zatiaľ čo l-glyceraldehyd prelomil rovinu polarizovaného svetla vľavo (-) a má rotáciu [α] D, pri 25 ° C, od -8.7 °.
Chirálny uhlík glyceraldehydu je uhlík 2 (C-2), čo je sekundárny alkohol. Fischerova projekcia predstavuje hydroxylovú skupinu (-OH) d-glyceraldehydu vpravo a OH-skupinu ľavého L-glyce.
Dihydroxyacetón nemá chirálne uhlíky a nemá enantiomérne formy. Pridanie hydroxymetylénovej skupiny (-choh) k glyceraldehydu alebo dihydroxyacetónu umožňuje vytvorenie nového chirálneho centra. V dôsledku toho je cukor tetrosa, pretože má štyri uhlíky.
Pridanie skupiny -cah do Terosa vytvára nové chirálne centrum. Tvorený cukor je pentóza. Skupiny -môžu byť naďalej pridané do maximálne desiatich uhlíkov.
Môže vám slúžiť: fylogéniaFunkcie v organizme
Triosy ako sprostredkovatelia v glykolýze, glukoneogenéze a dráhe fosfátu pentosa
Glykolýza pozostáva z prasknutia molekuly glukózy v dvoch pyruvátových molekulách na výrobu energie. Táto trasa znamená dve fázy: 1) prípravná fáza alebo spotreba energie; 2) Fáza tvorby energie. Prvý je ten, ktorý produkuje triosy.
V prvej fáze sa energetický obsah bez glukózy zvyšuje tvorbou fosfoésters. V tejto fáze je adenozín triffosfát (ATP) darcom fosfátu. Táto fáza vyvrcholila konverziou fruktózy fosfoéstera 1.6-bifosfátu (F1.6bp) v dvoch triosasových fosforečnanoch, glyceraldehyd 3-fosfátu (GA3P) a dihydroxyacetón-fosfát (DHAP) (DHAP) (DHAP) (DHAP) (DHAP).
Glukoneogenéza je biosyntéza glukózy z pyruvátu a iných sprostredkovateľov. Používajte všetky glykolýzové enzýmy, ktoré katalyzujú reakcie, ktorých biochemická štandardná variácia energie je v rovnováhe (ΔGº '~ 0). Z tohto.
Cesta pentózy fosfátu pozostáva z dvoch štádií: oxidačnej fázy glukózy-6-fosfátu a ďalšia tvorba NADPH a RibosA-5-fosfátu. V druhej fáze sa rebóza 5-fosfátu premieňa na sprostredkovateľov glykolýzy, F1.6bp a GA3P.
Triosas a cyklus Calvin
Fotosyntéza je rozdelená do dvoch etáp. V prvom sa vyskytujú reakcie závislé od svetla, ktoré produkujú NADPH a ATP. Tieto látky sa používajú v druhej, v ktorej dochádza k fixácii oxidu uhličitého a hexózy z Triosas cez cestu známu ako Calvin Cycle.
V Calvinovom cykle 1,5-bifary-oxygenáza (Rubisco) ribulózový enzým katalyzuje kovalentný zväzok CO2 Do pentózy Ribulosa 1,5-bifosfát a zlomí nestabilný sprostredkovateľ šiestich atómov uhlíka v dvoch molekulách troch atómov uhlíka: 3-fosfoglycelera.
Môže vám slúžiť: oligosacharidy: charakteristiky, zloženie, funkcie, typyProstredníctvom enzymatických reakcií, ktoré zahŕňajú fosforyláciu a redukciu 3-fosfoglycelera, s použitím ATP a NADP, sa vyskytuje GA3P. Tento metabolit sa premieňa na 1,6-bifosfátovú fruktózu (F1.6bp) metabolickou dráhou podobnou glukoneogenéze.
Pôsobením fosfatázy sa F1.6bp premení na fruktózu-6-fosfát. Potom fosfhexóza izomázy produkuje glukózu 6-fosfát (GLC6P). Nakoniec epicherasa prevádza GLC6P na glukózu 1-fosfát, ktorý slúži na biosyntézu škrobu.
Triosy a lipidy biologických a adipocytových membrán
GA3P a DHAP môžu tvoriť fosfát glycerol, ktorý je nevyhnutným metabolitom pre biosyntézu triacylglycerolov a glycerolipidov. Dôvodom je skutočnosť, že oba triosas fosfát môže byť vzájomne prepojený reakciou katalyzovaným pomocou triosa izomerázového fosfátu, ktorý udržiava obe triosy v rovnováhe.
Glycerol-fosfát enzým dehydrogenáza katalyzuje oxidovú redukciu, v ktorej NADH daruje pár elektrónov DHAP za vzniku 3-fosfátového glycerolu a NAD+. L-glycerol 3-fosfát je súčasťou kostry fosfolipidov, ktoré sú štrukturálnou súčasťou biologických membrán.
Glycerol je prozačný, chýba mu asymetrické uhlíky, ale keď jeden z jeho dvoch primárnych alkoholov tvorí fosfoester, môže sa správne nazývať l-glycerol 3-fosfát alebo d-glycerol 3-fosfátu.
Glyceofosfolipidy sa tiež nazývajú fosfoglyceridy, ktoré sa nazývajú ako kyselina fosfatidová. Fosfoglyceridy môžu tvoriť fosfoacylglycelály vytvorením esterových väzieb s dvoma mastnými kyselinami. V tomto prípade je výsledným produktom 1,2-fosfodiacylglycerol, ktorý je dôležitou súčasťou membrán.
Glyceofosfáza katalyzuje hydrolýzu skupiny 3-fosfátového glycerolu, produkuje glycerol viac fosfátu. Glycerol môže slúžiť ako počiatočný metabolit pre biosyntézu triacylglyceridov, ktoré sú bežné v adipocytoch.
Môže vám slúžiť: koagulázový test: nadácia, postup a použitieTriosy a membrány archeobaktérií
Podobne ako eubaktérie a eukaryoty, 3-fosfát glycerol sa tvorí z Triosas fosforečnanu (GA3P a DHAP). Existujú však rozdiely: prvá je, že 3-fosfát glycerol v archeobaktériách membrán je konfigurácia L, zatiaľ čo v eubaktériách a eukaryotických membránach je konfigurácia d.
Druhým rozdielom je, že archeobaktériové membrány tvoria esterové väzby s dvoma dlhými uhľovodíkovými reťazcami izoprenoidných skupín, zatiaľ čo v eubaktériách a eukaryotoch glycerolové formy esterových väzieb (1,2-diacilglycerol) s dvoma uhľovodíkovými reťazcami mastných kyselín.
Tretím rozdielom je to, že v membránach archeobaktérií sa náhrady vo fosforečnanskom a 3-fosfátovom glycerolu líšia od eubaktérií a eukaryotov. Napríklad skupina fosfátov je spojená s disacharidom α-Glucopiranso- (1®2)-p-Galathofuranosa.
Odkazy
- Cui, s. W. 2005. Potravinové uhľohydráty: chémia, fyzikálne vlastnosti a aplikácie. CRC Press, Boca Raton.
- Cock, P., Mäkinen, K, Honkala a., Saag, m., Kennepohl, e., Eapen, a. 2016. Erytritol je účinnejší ako xylitol a sorbitol pri riadení koncových bodov zdravia orálnych zdravotníctva. Medzinárodný denník zubného lekárstva.
- Nelson, D. L., Cox, m. M. 2017. Lehninger princípy biochémie. W. H. Freeman, New York.
- Sinnott, m. L. 2007. Sacharidová chémia a biochémia štruktúra a mechanizmus. Royal Society of Chemistry, Cambridge.
- Stick, r. Vložka., Williams, s. J. 2009. Sacharidy: základné molekuly života. Elsevier, Amsterdam.
- Voet, D., Voet, J. G., Pratt, C. W. 2008. Základy biochémie - život na molekulárnej úrovni. Wiley, Hoboken.
- « Charakteristiky aldózy, typy, počet uhlíkov
- Fluoxetín (Prozac) Mechanizmus účinku a vedľajších účinkov »