

Štruktúra uracilu, funkcie, vlastnosti, syntéza
- 4199
- 881
- Gabriel Bahna
On Uracil Je to báza dusíka pyrimidínu, ktorá sa nachádza v kyseline ribonukleovej (RNA). Toto je jedna z charakteristík, ktorá odlišuje RNA od kyseliny deoxyribonukleovej (DNA), pretože táto má Timinu namiesto uracilu Timinu. Obe látky, Uracil a Timina, sa líšia iba v tom, že druhá má metylovú skupinu.
Z evolučného hľadiska bolo navrhnuté, že RNA bola prvou molekulou, ktorá ukladala genetické informácie a pracovala ako katalyzátor v bunkách pred DNA a enzýmami. Z tohto dôvodu sa predpokladá, že Uracil mal kľúčovú úlohu vo vývoji života.
Zdroj: Kemikungen [verejná doména] U živých bytostí sa Uracil nenachádza voľne, ale bežne monofosfátových nukleotidov (UMP), difosfátu (UDP) a trfosfátu (UTP). Tieto uracilové nukleotidy majú rôzne funkcie, ako je biosyntéza RNA a glykogén, izomérna interkonverzia cukrov a regulácie syntézy syntézy synthamín.
[TOC]
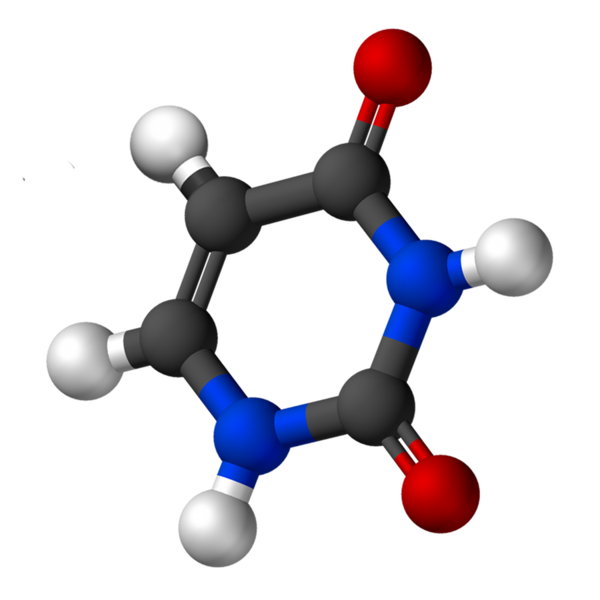
Štruktúra
Uracil, nazývaný 2,4-dioximiridín, má empirický vzorec C4H4N2Ani2, ktorej molekulová hmotnosť je 112,09 g/mol a je čistená ako biely prášok.
Štruktúra uridínu je heterocyklický kruh so štyrmi atómami uhlíka a dvoma dusíkmi, s dvojitými alternatívnymi väzbami. Je to planár.
Má rozpustnosť 50 mg/ml, 25 ° C, v 1M hydroxidu sodného a PKA medzi 7.9 a 8.2. Vlnová dĺžka, kde dochádza k jej maximálnej absorbancii (ʎMaximálny) Je medzi 258 a 260 nm.
Biosyntéza
Existuje spoločná cesta pre biosyntézu pyrimidínových nukleotidov (uracil a cytokín). Prvým krokom je biosyntéza fosforečnanu karbamilu od CO2 a NH4+, ktorý je katalyzovaný fosfátom syntetázy karbamil syntetázy.
Pyrimidín je postavený z karboilového fosfátu a aspartátu. Obe látky reagujú a tvoria N-CarbamoMoAspartato, reakcia katalyzovaná transcabamoilizou aspartát (ATCASA). Uzatvorenie pyrimidínového kruhu môže byť dehydratované dihydroottázou a vytvára L-dihydrotatoát.
L-dihydrotoát sa oxiduje a mení sa na orotát; Elektrónový akceptor je NAD+. Je to reakcia katalyzovaná dihydroorotátovou dehydrogenázou. Ďalší krok spočíva v prenose fosforibosylovej skupiny z fosforibozyl pyrofosfátu (PRPP) na orotáciu. Gradiditato forma (WPO) a anorganický piroffosfat (PPI), katalyzovaný fosforibosylou -orotátovou prenosázou.
Posledný krok pozostáva z dekarboxylácie pyrimidínového kruhu skupiny (WPO). Forma uridilaato (uridin-5'-monofosfát, ump), ktorý je katalyzovaný dekarboxylázou.
Potom sa prostredníctvom účasti kinázy prenáša fosfátová skupina z ATP do UMP, ktorá tvorí UDP (uridin-5'-difosfát). Opakuje sa, vytvára UTP (uridin-5'-triffosfát).
Môže vám slúžiť: vetvy biológie a čo študujúRegulácia biosyntézy
V baktériách dochádza k regulácii biosyntézy pyrimidínu prostredníctvom negatívnej spätnej väzby na úrovni aspartátu Transcabamoilasa (ATCASA).
Tento enzým je inhibovaný CTP (citidín-5'-trofosfát), ktorý je konečným produktom biosyntézy dráhy pyrimidínov. Atcas.
U zvierat sa dochádza k regulácii biosyntézy pyrimidínu prostredníctvom negatívnej spätnej väzby na úrovni dvoch enzýmov: 1) fosfát syntázy II karbamilu, ktorý je inhibovaný UTP a aktivovaný pomocou ATP a PRPP; a 2) OMP Discarboxyláza, ktorá je inhibovaná produktom reakcie, ktorú sama katalyzuje, UMP. Rýchlosť biosyntézy OMP sa líši v závislosti od dostupnosti PRPP.
Funkcia v biosyntéze RNA
Uracil je prítomný vo všetkých typoch RNA, ako je napríklad Messenger RNA (MNA), prenosová RNA (ARNT) a ribozomálna RNA (RNA). Biosyntéza týchto molekúl sa vyskytuje procesom nazývaným transkripcia.
Počas prepisu sa informácie obsiahnuté v DNA skopírujú v RNA cez Pon polymerázovú RNA. Reverzný proces, v ktorom sa informácie obsiahnuté v RNA skopírujú v DNA, sa vyskytuje v niektorých vírusoch a rastlinách prostredníctvom reverznej transkriptázy.
Biosyntéza RNA potrebuje skúšobné nukleozidy (NTP), menovite: tryfosfátový uridín (UTP), Tryposfát cytidín (CTP), adenínový pokus adenín (ATP) a tryfosfát Guanine (GTP). Reakcia je:
(RNA)n odpad + NTP -> (RNA)N+1 odpad + PPI
Hydrolýza anorganického pyrofosforečnanu (PPI) poskytuje energiu na biosyntézu RNA.
Funkcia v biosyntéze cukru
Estery cukru sú v živých organizmoch veľmi bežné. Niektoré z týchto esterov sú diphospas nukleozidových esterov, ako je napríklad UDP-cukr, ktoré sú v bunkách veľmi hojné. UDP-sugar zahŕňa biosyntézu disacharidov, oligosacharidov a polysacharidov.
V rastlinách sa biosyntéza sacharózy koná dvoma spôsobmi: hlavný a sekundárny.
Hlavná trasa spočíva v prenose d-glukózy z UDP-D-glukózy do d-plukingu za vzniku sacharózy a UDP. Sekundárna cesta obsahuje dva kroky: Začína sa uDP-D-glukózou a fruktóza-6-fosfát a kulminuje tvorbou sacharózy a fosfátu.
V prsných žľazach sa biosyntéza laktózy uskutočňuje z UDP-D-Galaktózy a glukózy.
Môže vám slúžiť: pobočky biochémieV rastlinách sa biosyntéza celulózy vykonáva kontinuálnou kondenzáciou beta-d-glykozylového odpadu, od UDP-glukózy po neredukčný koniec rastúceho polyglukózového reťazca. Podobne biosyntéza amylózy a amylopectínu vyžaduje UDP-glukózu ako substrát darcu glukózy do rastúceho reťazca.
U zvierat sa na biosyntézu glykogénu používajú UDP-glukóza aj ADP-glukóza. Podobne biosyntéza sulfátu chondroitínu vyžaduje UDP-xylose, UDP-galaktózu a UDP-glukuronato.
Funkcia v izomérnej vzájomnej vzájomnej interkonverzii cukrov
Konverzia galaktózy na sprostredkovateľ glykolýzy sa uskutočňuje cez cestu Leloir Road. Jedným z krokov tejto trasy je katalyzovaný enzýmom UDP-galaktózy-4-epimerázy, ktorý uľahčuje interkonverziu UDP-galaktózy do UDP-glukózy.
Funkcia v biosyntéze glykoproteínov
Počas biosyntézy glykoproteínov proteíny prechádzajú cez CIS, stredné a trans vrece Golgiho aparátu.
Každá z týchto tašiek má sadu enzýmov, ktoré spracúvajú glykoproteíny. Monoméry cukru, ako je glukóza a galaktóza.
Nukleotidové hexózy sa transportujú do nádrží Golgi Antiporte. UDP-galaktóza (UDP-GAL) a UDP-N-Acetylgalactosamina (UDP-GALNAC) vstupujú do nádrží z cytosolu pomocou výmeny pomocou UMP.
V nádrži Golgi fosfatázy hydrolyzuje fosfátovú skupinu formy UDP a UMP a PI. UDP pochádza z reakcií katalyzovaných galaktozyltransferázou a N-acetylgalaktosamiltransferáza. UMP vytvorená fosfatázou slúži na výmenu nukleotidových hexóz.
Funkcia v regulácii syntázy
Mechanizmom regulácie glutamín syntázy je kovalentná modifikácia, ktorá pozostáva z adenilatácie, ktorá je neaktívna a bok, ktorá ho aktívne aktívne. Táto kovalentná modifikácia je reverzibilná a katalyzovaná adenyltransferázou.
Aktivita adenyltransferázy je modulovaná spojením proteínu PII, ktorý je regulovaný kovalentnou modifikáciou, uridinilatáciou.
Uridililation aj ofset sa vykonávajú uridililtransferáza. V tomto enzýme je určená určená aktivita spôsobená glutamínom a fosfátom a je aktivovaná spojením alfa-zotoglutarátu a ATP na PII.
Funkcia v vydaní RNA
Niektoré RNM sú upravené pred prekladom. V niektorých eukaryotických organizmoch, ako napríklad Trypanosoma brucei, Existuje vydanie RNA transkriptu génu podjednotky II cytochrómu oxidázy. Deje sa to vložením odpadu z uracilu, čo je reakcia katalyzovaná terminálnou uridiltransferázou.
Môže vám slúžiť: Curl: Zloženie, časti, dôležitosťSprievodca RNA, dopĺňajúca upraveného produktu, pôsobí ako mierny pre proces úpravy. Páry báz vytvorených medzi počiatočným prepisom a vodiacou RNA naznačuje páry báz G = u, ktoré nie sú Watsonov rekord a sú bežné v RNA.
Biosyntéza UDP-glukóza
Vo fyziologických podmienkach je biosyntéza glykogénu z glukózy-1-fosfátu termodynamicky nemožná (pozitívny AG). Z tohto. Tento G1P a UTP kombinujú reakciu za vzniku glukózového difosfátu urridínu (UDP-glukóza alebo UDPG).
Reakcia je katalyzovaná pyrofosforylálom UDP-glukózy a je nasledovná:
G1p + utp -> udp -glucosa + 2pi.
Variácia voľnej energie Gibbs v tomto kroku je veľká a negatívna (-33,5 kJ/mol). Počas kyslíkovej reakcie G1P útočí na atóm alfa alfa alfa fosforu UDP-glukózy a anorganického pyrofosfátu (PPI). Potom sa PPI hydrolyzuje anorganickými pyrofosfátami, ktorých hydrolýza je tá, ktorá riadi všeobecnú reakciu.
UDP-glukóza je látka „s vysokou energiou“. Umožňuje tvoriť glykozidické väzby medzi zvyškom glukózy a zvyšujúcim sa polysacharidovým reťazcom. Rovnaký princíp energetiky je použiteľný na reakcie, na ktorých sa zúčastňuje cukru UDP, ako je biosyntéza disacharidov, oligosacharidov a glykoproteínov.
Uracil DNA glycosilalasa
Existujú lézie DNA, ktoré sa vyskytujú spontánne. Jednou z týchto lézií je sprontán. V tomto prípade sa oprava uskutoční v dôsledku modifikovanej bázy DNA pomocou enzýmu nazývaného Uracil DNA glycosilalasa.
Enzým uracil DNA glykozilasa eliminuje poškodený cytokín (uracil), ktorý produkuje deoxyribózový zvyšok, ktorý nemá dusíkovú bázu, nazývanú miesto AP (miesto apurinika-apirimidínu).
Potom enzým Endonuklease AP urobí strih vo fosfodiesterovom kostre v mieste AP, čím sa eliminuje zvyšok cukru a fosfátu. DNA polymeráza I obnovuje poškodený prameň.
Odkazy
- Bohinski, r. 1991. Biochémia. Addison-Wesley Iberoamericana, Wilmington, Delaware.
- Devlin, T.M. 2000. Biochémia. Redakčné Reverté, Barcelona.
- Ubytovňa, h., Berk, a., Zipurski, s.L., Matsudaria, P., Baltimore, D., Darnell, J. 2003. Bunková a molekulárna biológia. Redakcia Medica Panamericana, Buenos Aires, Bogota, Caracas, Madrid, Mexiko, Sāo Paulo.
- Nelson, D. L., Cox, m. M. 2008. Lehninger-Principles of Biochemistry. W.H. Freeman, New York.
- Voet, D. a Voet, J. 2004. Biochémia. John Wiley a Sons, USA.
- « Vzorec marginálnych nákladov, výpočet a cvičenia
- Charakteristiky pyrimidínov, štruktúra, funkcie »