

Štruktúra rastového hormónu (somatotropín), funkcie
- 4835
- 1249
- Adrián Fajnor
Ten Somatotropín (STH) U Rastový hormón (GH) je relatívne malý proteín produkovaný na úrovni adenohypofýzy a podieľa sa na vývojových procesoch, pozdĺžnom raste organizmu a kontrole niekoľkých metabolických procesov.
Je to ne -glaro hormón. Glandotropas hormóny hypofýzy sa zhromažďujú.
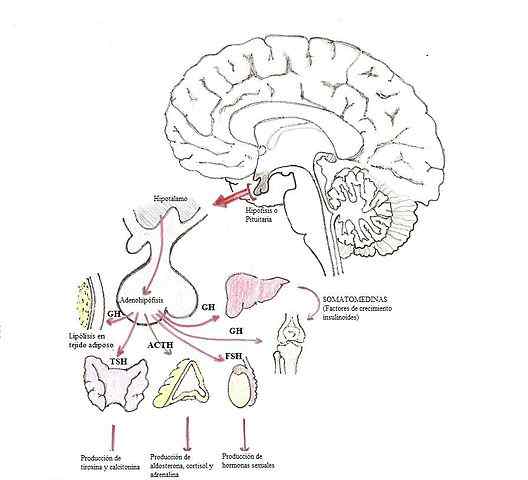
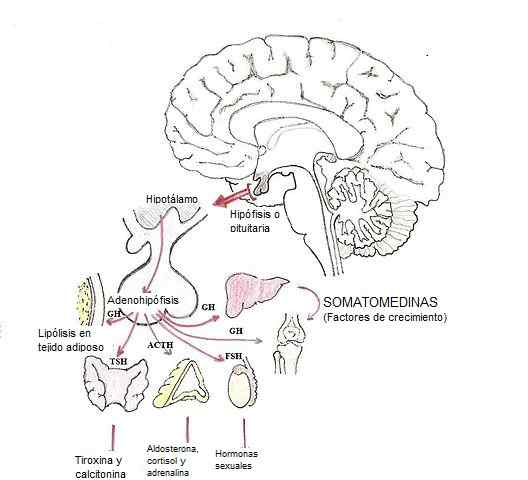 Modifikácia hormónov v hypofýze (Zdroj: Dubaele / Public Domain, Via Wikimedia Commons)
Modifikácia hormónov v hypofýze (Zdroj: Dubaele / Public Domain, Via Wikimedia Commons) Hormóny glandotropov sú napríklad adrenokortopín (ACTH), gonadotropíny (FSH a LH) a hormón stimulujúci štítna žľaza (TSH).
Non -glandropas hormóny, na druhej strane a medzi ktorými je zahrnutý prolaktín a rastový hormón.
[TOC]
Štruktúra
Rastový hormón je relatívne malý proteín, ktorý sa vyskytuje v niekoľkých izoformách. Hlavná izoforma sa skladá z približne 191 aminokyselín, má molekulovú hmotnosť 22 kDa a pochádza z dlhšieho prekurzora peptidu (pred GH) 28 kDa, ktorý sa tiež vylučuje, ale chýba to fyziologické funkcie.
Zdá sa, že vo svojej štruktúre je somatotropín homologovaný s prolaktínom a chorionickým somatomamamamototropínom (CS), ktorý bol vyrobený v placente. Taká je podobnosť, ktorá sa považuje za to, že títo traja tvoria hormonálnu rodinu.
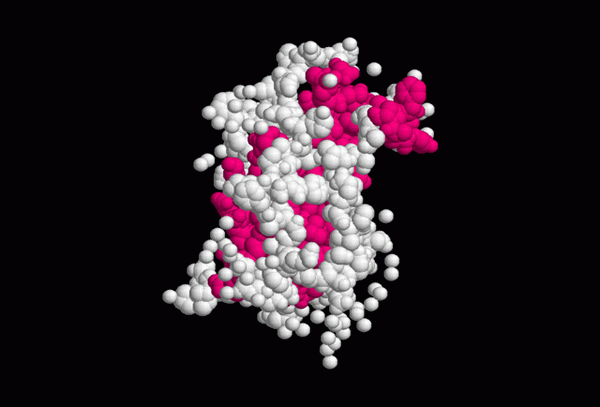 Približná štruktúra rastového hormónu (zdroj: роман бекер, cez Wikimedia Commons)
Približná štruktúra rastového hormónu (zdroj: роман бекер, cez Wikimedia Commons) Sekundárna štruktúra somatotropínu ukazuje 4 vrtule alfa stabilizované dvoma disulfidovými mostmi, ktorých konfigurácia je potrebná na interakciu hormónu s jeho prijímačom.
Fakt vo vzťahu k štruktúre a hodnosti zdôraznenia predstavuje skutočnosť, že napriek skutočnosti, že rastové hormóny rôznych druhov majú pozoruhodné podobnosti s človekom, iba druhým a primátmi, ktoré majú účinky významné u ľudí.
Funkcia
Somatotropínové funkcie sa zvyčajne opisujú ako funkcie spojené s vývojom a rastom organizmu. Tiež spojené s metabolizmom, ktoré zahŕňajú modifikácie metabolizmu lipidov a glukózy podporované hormónom.
Rastové funkcie sa však dajú považovať aj za metabolické, pretože naznačujú anabolické funkcie spojené so syntézou proteínov, ktoré nevylučujú niektoré iné funkcie bez priameho spojenia s metabolizmom, ako je proliferácia buniek, ako je bunková proliferácia.
Niektoré z funkcií alebo akcií nasadených somatotropínom sú vykonávané týmto hormónom priamo nad svojimi bielymi tkanivami, ale mnohé z nich sú vykonávané niektorými ďalšími látkami, ktorých syntéza a uvoľňovanie sú stimulované rastovým hormónom.
Syntéza IGF
Prvým riadkom somatotropínového pôsobenia je presne syntéza týchto látok, ktoré sú známe ako rastové faktory podobné inzulínu (IGF), z ktorých typy 1 a 2 boli identifikované. Sú označené ako IGF1 (najdôležitejšie) a IGF2, podľa svojej skratky v angličtine.
Tieto faktory boli spočiatku známe a sú stále označované ako mediátory somatotropínu alebo somatomedínov C (IGF1) a A (IGF2) alebo tiež ako aktivita podobná inzulínu bez stlačenia inzulínu (NSILA). Sú syntetizované mnohými typmi buniek, ale vyskytujú sa hlavne v pečeni.
Akcie STH a IGF1 sú veľmi rozmanité. Niektorí cvičia každú z týchto látok nezávisle, niekedy v spojení a synergicky a inokedy konajú antagonicky.
Môže vám slúžiť: elektroforéza: základ, technika, na čo je, príkladyIndukcia rastu
Toto je jedna z najdôležitejších akcií podporovaných somatotropínom, ale vykonaná v spojení s IGF1. Aj keď obidve vyvolávajú rast mnohých telových tkanív, ich najvýznamnejší účinok sa vyvíja na rast kostry.
Tento konečný výsledok je produkovaný niekoľkými účinkami vyvolanými hormónom a IGF1. Zahŕňajú zvýšenie ložiska proteínov chondrocytickými a osteogénnymi bunkami, najvyššiu rýchlosť reprodukcie týchto buniek a konverzia chondrocytov na osteogénne bunky; To všetko vedie k novému vkladu kostí.
Počas rastu a rozvoja organizmu a pred uzavretím kostných epifýz sa nová chrupavka ukladá v epifýnoch, po ktorých nasleduje jeho premena na novú kosť, ktorá predlžuje diafýzu a epifýzy sa oddeľujú.
Progresívna konzumácia epifyzovej chrupavky ju vyčerpá a kosť nemôže naďalej rásť. Na konci dospievania sa diafýza a epifýzy a rast dlhej dĺžky kostí spomaľujú na každý koniec a nakoniec sa zastaví.
Druhý mechanizmus môže vysvetliť zvýšenie hrúbky kostí. Osteoblasty periosteium ukladajú novú kosť na starú a osteoklasty eliminujú starú kosť. Ak rýchlosť depozície prekročí elimináciu, zvyšuje sa hrúbka.
Keďže rastový hormón cvičí intenzívnu stimuláciu na osteoblastoch, môže sa naďalej zvyšovať hrúbka kostí, hoci jej dĺžka už nie je modifikovaná v dôsledku uzavretia epifýz.
Propagácia ložiska proteínov v tkanivách
Tento účinok sa dá dosiahnuť rôznymi mechanizmami: zvýšenie transportu aminokyselín cez bunkové membrány, zvýšenie translácie RNA na hladine ribozómov, zvýšenie transkripcie DNA na RNA v jadre a zníženie proteínov a aminokyselín katabolizmus.
Ostatné metabolické akcie
V mastnom tkanive rastový hormón podporuje lipolýzu a uvoľňovanie mastných kyselín do obehového torrentu, ktorý zvyšuje jeho koncentráciu v telesných tekutinách. Zároveň uprednostňuje vo všetkých tkanivách konverziu mastných kyselín v acetylovom koenzýme A a jeho použitie ako zdroj energie.
Stimulácia použitia tukov spolu s akumuláciou proteínov v dôsledku ich anabolického účinku vedie k zvýšeniu chudého tkaniva.
Zvýšenie mobilizácie tukov môže byť také vysoké, že pečeň produkuje veľké množstvo kyseliny acetooctovej, ktorá vedie k ketóze a mastná pečeň.
V súvislosti s metabolizmom uhľohydrátov zahŕňajú účinky somatotropínu zníženie zberu glukózy v tukových a svalových svalových tkanivách, zvýšená produkcia pečene glukózy a zvýšená sekrécia inzulínu.
Všetky tieto účinky sa nazývajú diabetogény a sekrécia s vysokým rastovým hormónom môže reprodukovať metabolické zmeny, ktoré sprevádzajú ne -inzulín -závislý diabetes typu II.
Iné funkcie
Anabolické a mitogénne účinky GH a IGF1 sa prejavujú aj v raste a funkcii srdca, pečeni, slezine, štítnej žľaze, týmusu a jazyka. Hormón môže prispieť k zahusťovaniu kože, k stimulácii potných žliaz a rastu vlasov.
V obličkách sa miera glomerulárnej filtrácie a zvýšenie syntézy kalcitriolu, čo uprednostňuje nielen rast, ale aj mineralizáciu kostnej kosti, ale aj kostnú mineralizáciu. Podporuje tiež erytropoézu a syntézu fibrinogénu a imunitnú odpoveď stimuláciou T lymfocytov a makrofágov.
Môže vám slúžiť: epitelové tkanivo: Charakteristiky, popis, funkcie, typyReceptory pre somatotropín
Somatotropínové účinky, vrátane podpory syntézy rastových faktorov podobných inzulínu, sú sprostredkované prostredníctvom ich spojenia so špecifickými receptormi exprimovanými v membránach bielych buniek.
Existujú dve formy týchto receptorov, z ktorých druhá je krátky (skrátený) variant prvého; Skrátená forma, ktorá inhibuje funkciu dlhého prijímača a že ak by sa nadmerne exprimovala, vyvolala by tkanivovú necitlivosť na hormón.
Dlhý prijímač sa skladá zo 638 aminokyselín a má extracelulárnu doménu 250, alfa transmembraálna doména asi 38 a intracelulárna doména 350 aminokyselín. Každá molekula somatotropínu končí spojením dvoch molekúl prijímača a produkuje to, čo sa nazýva dimerizácia receptora.
Táto dimerizácia aktivuje proteín JAK2 kinázy umiestnený na intracelulárnych koncoch každého monoméru prijímača a tieto aktívne fosforylácie kinázy Fosforyl ostatné substráty, ako je STAT5 a samotný somatotropínový receptor.
Fosforylované molekuly STAT5 tiež trpia dimerizáciou, vďaka ktorej sú veľmi presné regulátory génovej expresie a syntézy proteínov.
Výroba
Somatotropín je syntetizovaný na úrovni adenohypofýzových somatotropasových buniek. Tieto bunky sú intenzívne farbené kyslými látkami, takže sa tiež nazývajú acidofily. Sú spolu najhojnejšou bunkovou skupinou žľazy, pretože predstavujú 50% z celkového počtu 5 rôznych typov.
V dlhom ramene ľudského chromozómu 17, ktorý kóduje rôzne izoformy rastového hormónu a ľudského chorionického somatomamotototropínu (HCS), je genetický komplex 5 génov.
Jedným z nich je normálny HGH-N, ktorý kóduje najhojnejšiu formu ľudského rastového hormónu, ktorý je vyššie uvedeným 22 kDa a predstavuje 75% celkového cirkulujúceho rastového hormónu.
Jeho Messenger RNA je vystavená „Strih a zostrihanie“ Na výrobu menšej formy hormónu, 20 kDa, ktorá nemá odpad z aminokyselín od 32 do 46 a predstavuje 10%.
Druhý gén (HGH-V) je exprimovaný hlavne v placente a kóduje variantnú formu HGH, ktorej iba významné množstvá sa objavujú v obehu počas tehotenstva. Ostatné 3 gény kódujú izoformy somatomamototropínu človeka.
Prepustenie
Syntéza aj sekrécia alebo uvoľňovanie rastového hormónu sú regulované stimulujúcimi a inhibičnými faktormi takýchto funkcií.
Stimulujúce vplyvy
Medzi hlavné humorné vplyvy stimulujúce syntézu a sekréciu somatotropínu patria peptidy GHRH (rastový hormón uvoľňujúci hormón) a ghrelín) a ghrelín.
Rastový hormón oslobodzujúci hormón (GHRH) je hypotalamický peptid, ktorý existuje v dvoch variantoch 40 a 44 aminokyselín. Vedie v somatotropasových bunkách do syntézy AMPC a na aktiváciu špecifického transkripčného faktora PIT1 pre rastový hormón.
Ghrelín je endogén rastového hormónu rastového hormónu. Je to peptid asi 28 aminokyselín syntetizovaných na úrovni hypotalamu a žalúdka. Pôsobí synergicky s GHRH, ktorého uvoľnenie podporuje, pričom inhibuje somatostatín. Pôsobí prostredníctvom receptorov, ktoré aktivujú fosfolipázu C.
Niektoré metabolické parametre, ako je hypoglykémia, nízke hladiny voľných mastných kyselín v krvi a vysoké koncentrácie aminokyselín sú dôležitými stimulmi pre sekréciu rastového hormónu.
Môže vám slúžiť: 12 stupňov ľudského rozvoja a jeho charakteristikyĎalšími stimulačnými faktormi, ktoré sa počítajú, sú akútny stres, preťaženie tela, bolesť, pohlavné steroidy (puberta), dopamín, stimulácia a2 receptorov, acetylcholín, galanín, serotonín a p-endorfín.
Inhibičné vplyvy
Medzi nimi patrí somatostatín alebo hormón inhibítor uvoľňovania rastového hormónu (GHRIH) a negatívna spätná väzba.
Somatostatín je hypotalamický peptid 14 aminokyselín, ktorý inhibuje sekréciu, ale nie syntézu rastového hormónu. V gastrointestinálnom trakte je syntetizovaný dlhý variant, 28 aminokyselín. Oba varianty sa viažu na rovnaký prijímač a inhibujú syntézu cyklickej AMP.
Pokiaľ ide o negatívnu spätnú väzbu, GH uvoľnil prostredníctvom sebapozácavého pôsobenia svoje ďalšie prepustenie. IGF1 inhibuje hypotalamus do rastového hormónu rastového hormónu a stimuluje somatostatín, zatiaľ čo inhibuje syntézu GH v hypofýze.
Niektoré metabolické parametre, ako je hyperglykémia, vysoké plazmatické hladiny voľných mastných kyselín a nízkymi hladinami aminokyselín sú inhibítory sekrécie somatotropínu.
Inhibítory sú tiež studené, chronické stresy, adipozita, progesterón, hormóny štítnej žľazy, deficity alebo excesy kortizolu a stimulácia adrenergných receptorov p2 β2.
Dávka
Terapeutické použitie biosyntetizovaného rastového hormónu je indikované na liečbu utrpení, v ktorých bol odhalený nedostatok jeho sekrécie, v hypofýze trpaslíku a u detí s nízkym obsahom Turnerovho syndrómu.
Správa je vyrobená vo forme injekčného roztoku prestavaného z cesty, ktorá obsahuje lyofilizovanú so 40 U.Jo. biosyntetického hormónu a do ktorého sa pridávajú 2 ml spoločníci s roztokom chloridu sodného.
V nedostatku rastového hormónu u detí sa odporúča medzi 0,07 a 0,1 u.Jo./kg telesná hmotnosť denne. V Turnerovom syndróme 0,14 u.Jo./kg telesná hmotnosť denne. Pre nedostatok rastového hormónu u dospelých: 0,018 až 0,036 u.Jo./kg telesná hmotnosť denne.
Účinky
Terapeutické podávanie rastového hormónu môže byť sprevádzané niektorými nepriaznivými kolaterálnymi účinkami, ako je zjavná hypersenzitivita prostredníctvom zovšeobecnenej urtikárie, hypoglykémie nalačno, zápal v mieste injekcie a bolesti hlavy cestujúcich.
Vývoj určitej benígnej intrakraniálnej hypertenzie bol opísaný, častejší u detí a menej u dospelých.
Pokiaľ ide o metabolizmus uhľohydrátov, bol hlásený vývoj diabetes mellitus u pacientov, ktorí dostávali liečbu rastovým hormónom.
Vo vzťahu k systému kostrových svalov existujú dôkazy o zápalovej myozitíde s myalgiou a svalovou slabosťou, ktoré nie sú produkované hormónom, ale možno aj metacresolom používaným ako konzervatívny vo vzorci.
Boli hlásené prípady gynekomastie, anémie a akútnej pankreatitídy.
Odkazy
- Ganong WF: The Hypofýza, 25. vydanie. New York, McGraw-Hill Education, 2016.
- Guyton AC, Hall JE: Hypofýzy a ich kontrola hypotalamom, v Učebnica lekárskej fyziológie, 13. Ed, AC Guyton, JE Hall (eds). Philadelphia, Elsevier Inc., 2016.
- Lang F, Verrey F: hormón, v Physiologie des Menschen Mite patofysiologie, 31. ed, RF Schmidt a kol. (Eds). Heidelberg, Springer Medizin Verlag, 2010
- Voigt K: Endokrines System, v Physiologie, 6. vydanie; R Klinke a kol. (Eds). Stuttgart, Georg Thieme Verlag, 2010.
- Widmaier EP, RAPH H a Strang KT: Endokrinný systém. Hypotalamus a hypofýza, vo Vanderovej ľudskej fyziológii: mechanizmy funkcie tela, 13. vydanie; EP Windmaier a kol. (Eds). New York, McGraw-Hill, 2014.
- « Koncept aliterácie, charakteristiky a príklady
- Charakteristiky skupinovej komunikácie, prvky, príklady »