

Štruktúra, typy, funkcie stredných vlákien
- 901
- 106
- Tomáš Klapka
Ten stredné vlákna, Tiež známe v literatúre ako „ifs“ (z angličtiny Medziprodukcia) sú rodinou nerozpustných cytosolických vláknitých proteínov, ktoré sú prítomné vo všetkých mnohobunkových eukaryotických bunkách.
Sú súčasťou cytoskeletu, ktorý je intracelulárnou vláknitou sieťou, ktorá je zodpovedná hlavne za podporu bunkovej štruktúry a rôznych metabolických a fyziologických procesov, ako je transport vezikúl, pohyb a presídlenie buniek atď.
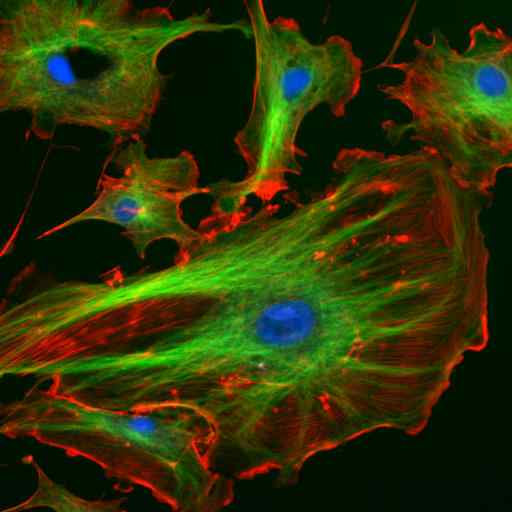
 Mikroskopia imunofluorescenciou dvoch proteínov medziproduktových filamentov astrocytov (Vimentina a GFAP) (Zdroj: Gerryshaw [CC By-SA 3.0 (https: // creativeCommons.Org/licencie/By-SA/3.0)] Via Wikimedia Commons)
Mikroskopia imunofluorescenciou dvoch proteínov medziproduktových filamentov astrocytov (Vimentina a GFAP) (Zdroj: Gerryshaw [CC By-SA 3.0 (https: // creativeCommons.Org/licencie/By-SA/3.0)] Via Wikimedia Commons) Spolu s mikrotubulami a mikrofilanciami sa stredné vlákna zúčastňujú na priestorovej organizácii intracelulárnych organel, v procesoch endocytózy a exocytózy a tiež v procesoch delenia buniek a medzibunkovej komunikácie.
Prvé stredne pokročilé vlákna, ktoré boli študované a opísané, boli keratíny, jeden z prvých typov proteínov, ktorých štruktúra bola analyzovaná X -BAY difrakciou v 30. rokoch 20. storočia.
Koncept stredných vlákien však zaviedol v 80. rokoch 20. storočia Lazarides, ktorý ich opísal ako „mechanických integrátorov bunkového priestoru“, ktorý sa vyznačuje ich nerozpustnosťou a schopnosťou reensovať sa In vitro Po jeho denaturácii.
Mnoho autorov sa považuje za „tlmiace“ prvky stresu pre živočíšne bunky, pretože sú flexibilnejšími vláknami ako mikrotubuly a mikrofilamenty. Nielenže sa nachádzajú v cytoskelete, ale sú tiež súčasťou nukleoesqueleta.
Na rozdiel od ostatných vláknitých komponentov cytoskeletu sa medziľahlé vlákna nezúčastňujú priamo na procesoch mobility buniek, ale pracujú na štrukturálnom udržiavaní a mechanickej odolnosti buniek.
[TOC]
Štruktúra
Zdroj: http: // rsb.Info.NIH.Gov/IJ/Images/[Public Domain] Medzi pokročilé vlákna majú približný priemer 10 nm, štrukturálnu charakteristiku, pomocou ktorej boli pomenované, pretože ich veľkosť je medzi veľkosťami zodpovedajúcimi myozínovým a aktínovým vláknom, ktoré sú okolo 25 nm a 7 nm, respektíve 7 nm.
Líšia sa štruktúrne od ostatných dvoch typov cytoskeletových vlákien, ktoré sú globulárnymi proteínovými polymérmi, v ktorých ich zložkové jednotky sú rôzne a-heliidálne vláknité proteíny s veľkou dĺžkou, ktoré sú navzájom zoskupené, aby vytvorili štruktúry vo forme lana alebo lana vo forme lana alebo lana vo forme lana alebo lana vo forme lana alebo lana vo forme lana alebo lana vo forme lana alebo lana sú.
Všetky proteíny, ktoré tvoria medziprodukčné vlákna, majú podobnú molekulárnu organizáciu, ktorá pozostáva z domény a-helicidej alebo „lana“, ktorá má rôzne množstvá segmentov „cievok“ rovnakej veľkosti.
Táto špirálová doména je lemovaná nekonečnou „hlavou“ a nehypoidálnym „chvostom“ na konci C-terminálneho konca a obidve sa menia vo veľkosti aj aminokyselinovej sekvencii.
V sekvencii týchto dvoch extrémov sú konsenzuálne dôvody, ktoré sú bežné pre 6 typov známych medziproduktov.
U stavovcov má „lano“ doména proteínu medziproduktov cytosolických vlákien asi 310 aminokyselinových odpadov, zatiaľ čo cytosolické proteíny bezstavovcov a jadrového laminátu majú dĺžku asi 350 aminokyselín.
Môže vám slúžiť: plazy: Čo sú, charakteristiky, klasifikácia, reprodukciazhromaždenie
Medziprodukčné vlákna sú štruktúry „samostatne zostavenia“, ktoré nemajú enzymatickú aktivitu, ktorá ich tiež odlišuje od ich cytoskeletálnych náprotivkov (mikrotubuly a mikrofilamenty).
Tieto štruktúry sa pôvodne zostavujú ako tetraméry vláknitých proteínov, ktoré ich tvoria pod vplyvom iba monovalentných katiónov.
Tieto tetraméry sú dlhé 62 nm a ich monoméry sú navzájom spojené laterálne, aby vytvorili „jednotky dĺžky“ (UFL, angličtiny Odborové filmy), Čo je známe ako fáza 1 zostavy, ktorá sa vyskytuje veľmi rýchlo.
UFL sú prekurzormi dlhých vlákien a keďže diméry, ktoré ich tvoria, sa navzájom viažu antiparallom a rozloženým spôsobom, tieto jednotky majú ústrednú doménu s dvoma lemujúcimi doménami, cez ktoré sa fáza 2 predĺže Z ďalších UFL je uvedený.
Počas toho, čo sa nazývalo fáza 3 zostavy, sa vyskytuje radiálne zhutnenie priemeru vlákien, ktoré vytvára zrelé medziprodukty s priemerom viac alebo menej 10 nm.
Funkcia
Funkcie medziľahlých vlákien značne závisia od typu zvažovaného bunky, av prípade zvierat (vrátane ľudí) je ich expresia regulovaná tkanivovo špecifickým spôsobom, takže záleží aj na type tkaniva ako v prípade štúdium.
Epitel, svaly, mezenchymálne a glile a neuróny majú rôzne typy vlákien, špecializované podľa funkcie buniek, ku ktorým patria.
Medzi týmito funkciami je najdôležitejšie štrukturálne udržiavanie buniek a rezistencia na rôzne mechanické napätie, pretože tieto štruktúry majú určitú pružnosť, ktorá im umožňuje tlmiť rôzne typy síl uložených na bunkách.
Typy stredných vlákien
Proteíny, ktoré tvoria stredné vlákna, patria do veľkej a heterogénnej rodiny vláknitých proteínov, ktoré sú chemicky odlišné, ale rozlišujú sa v šiestich triedach podľa ich homológie sekvencií (I, II, III, IV, V a VI).
Aj keď to nie je veľmi bežné, rôzne typy buniek, za veľmi konkrétnych podmienok (vývoj, transformácia buniek, rast atď.) Môžu exprimovať viac ako jednu triedu proteínov tvoriacich medziprodukty, ktoré tvoria proteíny
Trieda I a II stredných vlákien: Kyselina a základné keratíny
Keratíny predstavujú väčšinu proteínov stredných vlákien a u ľudí predstavujú viac ako tri štvrtiny medziproduktových vlákien.
Majú molekulové hmotnosti, ktoré sa pohybujú medzi 40 a 70 kDa a líšia sa od iných proteínov medziproduktu vlákna kvôli ich vysokému obsahu v glycíne a seríne.
Môže vám slúžiť: Hedgehogy: Charakteristiky, biotop, reprodukcia, jedloSú známe ako kyslé a základné keratíny pre svoje izoelektrické body, ktoré sú medzi 4.9 a 5.4 pre kyslé keratíny a medzi 6.1 a 7.8 pre základy.
V týchto dvoch triedach boli opísané okolo 30 proteínov a sú prítomné najmä v epitelových bunkách, kde oba typy proteínov „kolomerizujú“ a tvoria zlúčeniny vlákna.
Mnohé z prípadov I Keratíny medziľahlých vlákien sa nachádzajú v štruktúrach, ako sú vlasy, nechty, rohy, hroty a pazúry, zatiaľ čo trieda II je v cytosole najjemnejšia.
Trieda III stredných vlákien: proteíny typu demine/vimentin
Vodopády sú kyslý proteín 53 kDa, ktorý má v závislosti od stupňa fosforylácie rôzne varianty.
Niektorí autori tiež nazývali vlákna pádu ako „svalové stredné vlákna“, pretože ich prítomnosť je dosť obmedzená, hoci v malých množstvách všetky typy svalových buniek.
V myofibriloch sa pády nachádzajú v línii Z, takže sa predpokladá, že tento proteín prispieva k kontraktívnym funkciám svalových vlákien pri práci v spojení myofibrilov a plazmatickej membrány.
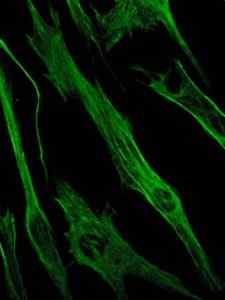 Fotografia zafarbenia vimentínového proteínu, proteínu medziproduktových vlákien epitelových a embryonálnych buniek (zdroj: Viktoriia Kosach [CC By-SA 4.0 (https: // creativeCommons.Org/licencie/By-SA/4.0)] Via Wikimedia Commons)
Fotografia zafarbenia vimentínového proteínu, proteínu medziproduktových vlákien epitelových a embryonálnych buniek (zdroj: Viktoriia Kosach [CC By-SA 4.0 (https: // creativeCommons.Org/licencie/By-SA/4.0)] Via Wikimedia Commons) Na druhej strane je vimentín proteín prítomný v mezenchymálnych bunkách. Medzi pokročilé vlákna tvorené týmto proteínom sú flexibilné a bolo stanovené, že odolávajú mnohým z konformačných zmien, ktoré sa vyskytujú počas bunkového cyklu.
Nachádza sa vo fibroblastoch, v bunkách hladkého svalstva, v bielych krvinkách a ďalších bunkách zvieracieho obehového systému.
Trieda IV stredných vlákien: Neurofilamentné proteíny
Tento druh stredných vlákien, tiež známy ako „neurofilamenty“, zahŕňa jeden zo základných štrukturálnych prvkov axónov a neuronálnych dendritov; Často sú spojené s mikrotubulami, ktoré tiež tvoria tieto štruktúry.
Neurofilamenty zvierat stavovcov boli izolované a určujú, že ide o 200, 150 a 68 kDa proteínový triplet, ktorý sa podieľa na montáži In vitro.
Líšia sa od iných stredne pokročilých vlákien, v ktorých majú bočné zbrane, ako napríklad „prílohy“, ktoré sú premietané z ich periférie a ktoré pracujú v interakcii medzi susednými vláknami a inými štruktúrami.
Gliové bunky produkujú špeciálny typ stredných vlákien známych ako gliové medziprodukty, ktoré sa štrukturálne líšia od neurofilamentov, pretože sú zložené z jediného 51 kDa proteínu a ktorý má rôzne fyzikálno -chemické vlastnosti.
Trieda V stredných vlákien: vlákna jadrových lamín
Všetky listy, ktoré sú súčasťou nukleoesqueleta, sú v skutočnosti stredné proteíny vlákna. Majú 60 a 75 kDa molekulovú hmotnosť a nachádzajú sa v jadrách všetkých eukaryotických buniek.
Môže vám slúžiť: Trichuris TrichiuraSú nevyhnutné pre vnútornú organizáciu jadrových oblastí a pre mnohé funkcie tejto základnej organely pre existenciu eukaryotov.
Trieda VI stredných vlákien: Nestiny
Tento typ stredných vlákien váži viac alebo menej 200 kDa a je prevažne v kmeňových bunkách centrálneho nervového systému. Sú vyjadrené počas vývoja neurónov.
Súvisiace patológie
U ľudských bytostí existuje viac chorôb, ktoré sa týkajú medziproduktových vlákien.
U niektorých typov rakoviny, ako sú napríklad malígne melanómy alebo karcinómy prsníka, vedie koexpresia medziproduktových vlákien vimentínu a keratínu k diferenciácii alebo vzájomnej interkonverzii epitelových a mezenchymálnych buniek.
Experimentálne sa preukázalo, že tento jav zvyšuje migračnú a invazívnu aktivitu rakovinových buniek, čo má dôležité dôsledky v metastatických procesoch charakteristických pre tento stav.
Eriksson a spolupracovníci (2009) Preskúmajú rôzne typy chorôb a ich vzťah so špecifickými mutáciami v génoch zapojených do tvorby šiestich typov medziproduktových vlákien.
Choroby súvisiace s kódujúcimi génovými mutáciami pre dva typy keratínu sú bullastová epidermolýza, epidermolytická hyperkeratóza, dystrofia rohovky, keratodermia a mnoho ďalších.
Medziprodukty typu III sa podieľajú na mnohých kardiomyopatiách a rôznych svalových chorobách súvisiacich hlavne s dystrofiami. Okrem toho sú tiež zodpovedné za dominantné katarakty a niektoré typy sklerózy.
Početné neurologické syndrómy a poruchy súvisia s vláknami typu IV, ako napríklad Parkinson Parkinson. Podobne sú genetické defekty vo vláknach typu V a VI zodpovedné za vývoj rôznych autozomálnych chorôb a súvisia s fungovaním bunkového jadra.
Príkladom sú syndróm Hutchinson-Gilford Progeria, Emery-Drifuss svalová dystrofia,.
Odkazy
- Anderton, b. H. (1981). Medziprodukčné filamles: Rodina homológnych štruktúr. Vestník svalového výskumu a pohyblivosti buniek, 2(2), 141-166.
- Eriksson, J. A., Pallari, h., Robert, D., Eriksson, J. A., Dechat, t., Úsmev, b.,... Goldman, r. D. (2009). Predstavujeme stredne pokročilé filamory: od objavovania po choroby. The Journal of Clinical Equesting, 119(7), 1763-1771.
- Fuchs, e., & Weber, K. (1994). Stredne pokročilé filamles: Štruktúra, dynamika, funkcia a choroba. Anu. Otáčať sa. Biochem., 63, 345-382.
- Hendrix, m. J. C., Seftor, e. Do., Chu a. W., Trevor, K. Tón., & Seftor, r. A. B. (Devätnásť deväťdesiat šiestich). Úloha medziproduktov pri migrácii, invázii a metastázovaní. Recenzie na rakovinu a metastázovanie, pätnásť(4), 507-525.
- Herrmann, h., & Aebi, u. (2004). Medziprodukty: Molekulárna štruktúra, mechanizmus zostavenia a integrácia do funkčne intracelulárnych skafoldov zreteľne. Ročné preskúmanie biochémie, 73(1), 749-789.
- Herrmann, h., & Aebi, u. (2016). Medziprodukčné filamles: Štruktúra a montáž. Perspektívy studeného jarného prístavu v biológii, 8, 1-22.
- McLean, i., & Lane, B. (Devätnásť deväťdesiatpäť). Stredne pokročilé filamory v Diew. Súčasný názor v bunkovej biológii, 7(1), 118-125.
- Steinert, P., & Roop, D. (1988). Molekulárna a bunková biológia medziproduktov. Ročné preskúmanie biochémie, 57(1), 593-625.
- Steinert, P., Jones, J., & Goldman, R. (1984). Medziprodukcia. The Journal of Cell Biology, 99(1), 1-6.
- « Gastrotrické charakteristiky, biotop, reprodukcia, jedlo
- Charakteristiky mäkkej mešity, taxonómia, biotop, použitie »