

Sfingolipidy Čo sú, charakteristiky, funkcie, syntéza
- 3219
- 630
- Denis Škriniar
Čo sú sfhylingolipidy?
Ten sfingolipidy Predstavujú jednu z troch veľkých lipidových rodín prítomných v biologických membránach. Rovnako ako glyceofosfolipidy a steroly, sú to amfipatické molekuly s hydrofilnou polárnou oblasťou a hydrofóbnou apolárnou oblasťou.
Prvýkrát ich opísal v roku 1884 Johann L. W. Thudichum, ktorý opísal tri sfingolipidy (sfingomyeline, mozog a mozgový mozg), ktoré patria do troch známych troch rôznych tried: fosfingolipidy, glykosfingolipidy neutrálne a kyslé.
Na rozdiel od glyceofosfolipidov nie sú sphyingolipidy postavené na molekule glycerolu 3-fosfátu ako hlavný kostra, ale sú to zlúčeniny odvodené od sfénu, aminoalkoholu s dlhým uhľovodíkovým reťazcom pripevnenom pomocou väzby amida.
AŠtruktúra sfingolipidov
Všetky sfingolipidy pochádzajú z L-serínu, ktorý sa kondenzuje mastnou kyselinou s dlhým reťazcom, aby sa vytvorila sfingoidná báza, tiež známa ako báza s dlhým reťazcom (LCB).
Najbežnejšími základňami sú sfinganín a sfinxín, ktoré sa navzájom líšia v prítomnosti dvojitej väzby previesť Medzi uhlíkmi 4 a 5 mastnej kyseliny sfinxínu.
Uhlíky 1, 2 a 3 sfinxínu sú štrukturálne analogické glycerofosfolipidom glycerolu glycerol. Keď väzby Amida viaže mastnú kyselinu na uhlík 2 sfhin.
Mastné kyseliny s dlhým reťazcom, ktoré tvoria hydrofóbne oblasti týchto lipidov, môžu byť veľmi rozmanité. Dĺžka sa pohybuje od 14 do 22 atómov uhlíka, ktoré môžu mať rôzne stupne saturácie, zvyčajne medzi uhlíkmi 4 a 5.
V pozíciách 4 alebo 6 môžu mať hydroxilli skupiny a dvojité väzby v iných pozíciách alebo dokonca dôsledkoch, ako sú metylové skupiny.
Charakteristika
Reťazce mastných kyselín spojené s väzbami Amida s ceramidmi sú bežne nasýtené a majú tendenciu mať väčšiu dĺžku ako tie, ktoré sa nachádzajú v glycerofosfolipidoch, čo sa zdá byť rozhodujúce pre biologickú aktivitu týchto.
Výraznou charakteristikou sfingolipidovej kostry je to, že môžu mať kladné čisté zaťaženie na neutrálne pH, čudné medzi lipidovými molekulami.
Však PkAminoskupina je nízka vzhľadom na jednoduchý amín, medzi 7 a 8, takže časť molekuly nie je naložená na fyziologické pH, čo by mohlo vysvetliť „voľný“ pohyb týchto medzi bilapami.
Tradičná klasifikácia sfingolipidov vyplýva z viacerých modifikácií, ktoré môže molekula ceramidu trpieť, najmä pokiaľ ide o substitúcie polárnych skupín hlavy.
Môže vám slúžiť: hepadnavírus: Charakteristiky, morfológia, liečbaFunkcia
Sfingolipidy sú nevyhnutné u zvierat, rastlín a húb, ako aj u niektorých prokaryotických organizmov a vírusov.
-Štrukturálne funkcie
Sfingolipidy modulujú fyzikálne vlastnosti membrán, vrátane ich plynulosti, hrúbky a zakrivenia. Modulárne tieto vlastnosti im tiež dávajú priamy vplyv na priestorovú organizáciu membránových proteínov.
V lipidových „plte“
V biologických membránach je možné detegovať dynamické mikro domény s nižšou plynulosťou tvorenou molekulami cholesterolu a sfingolipidov nazývané lipidové balzasy.
Tieto štruktúry sa vyskytujú prirodzene a udržiavajú úzky vzťah s komplexnými proteínmi, bunkovými povrchovými receptormi a signalizačnými proteínmi, transportérmi a iným proteínom s kotvami glykozylfosfatidylinitolu (GPI) (GPI).
-Signalizačné funkcie
Majú funkcie, ako sú signalizačné molekuly, ktoré pôsobia ako druhí poslovia alebo ako sekretované ligandy pre receptory bunkového povrchu.
Ako sekundárne poslovia sa môžu podieľať na regulácii homeostázy vápnika, rastu buniek, tumorogenézy a potlačenia apoptózy. Okrem toho aktivita mnohých integrálnych a periférnych membránových proteínov závisí od jej spojenia s sfingolipidmi.
Mnoho medzibunkových a bunkových interakcií s ich okolím závisí od expozície rôznych polárnych skupín sfingolipidov smerom k vonkajšej tvári plazmatickej membrány.
Spoja glykosfingolipidov a lektínov je rozhodujúci pre spojenie myelínu s axónmi, adhéziu neutrofilov do endotelu atď.
Podľa produktov jeho metabolizmu
Najdôležitejšími signalizačnými sfingolipidmi sú bázy s dlhým reťazcom alebo sfiny a ceramidy, ako aj ich fosforylované deriváty, ako napríklad 1-fosfát sfygosín.
Metabolizmus produktov mnohých sfingolipidov aktivuje alebo inhibuje viac bielych downstants (kinázové proteíny, fospatóza a ďalšie), ktoré kontrolujú bunkové správanie tak komplexné ako rast, diferenciácia a apoptóza.
-Ako membránové receptory
Niektoré patogény používajú gloysphingolipidy ako receptory na sprostredkovanie ich vstupu do hostiteľských buniek alebo na dodávku virulenčných faktorov im.
Ukázalo sa, že sfhyglyipidy sa podieľajú na viacerých bunkových udalostiach, ako je sekrécia, endocytóza, chemotaxia, neurotransmisia, angiogenéza a zápal.
Sú tiež zapojení do membránovej premávky, takže ovplyvňujú internalizáciu receptorov, poriadku, pohyb a fúziu sekrečných vezikúl v reakcii na rôzne podnety.
Sfingolipidové skupiny
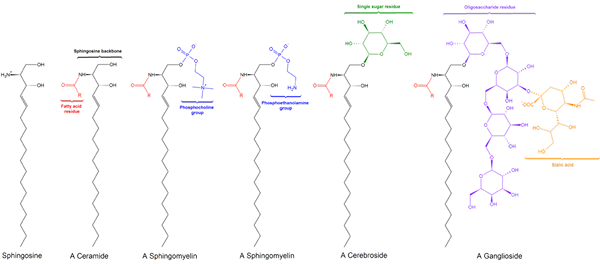
Existujú tri sfingolipidové podtriedy, všetky odvodené od ceramidu a ktoré sa navzájom líšia polárnymi skupinami, konkrétne: sfhylingomyelíny, glykolipidy a gangliazby.
Sfingomyielinas
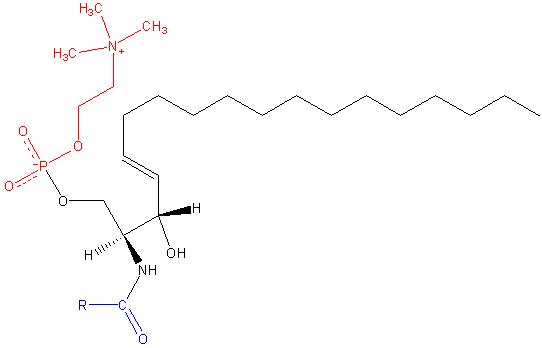 Sfingomileín. Čierna: sfinga. Červená: fosfokolín. Modrá: mastná kyselina.
Sfingomileín. Čierna: sfinga. Červená: fosfokolín. Modrá: mastná kyselina. Tieto obsahujú fosfokolín alebo fosfoetanolamín ako skupinu polárnej hlavy, takže sú klasifikované ako fosfolipidy spolu s glyceofosfolipidmi. Vyzerajú, samozrejme, fosfatidylcolins v trojrozmernej štruktúre a všeobecných vlastnostiach, pretože vo svojich polárnych hlavách nemajú žiadne poplatky.
Môže vám slúžiť: enolase: štruktúra, mechanizmus pôsobenia, funkcieSú prítomné v plazmatických membránach živočíšnych buniek a sú obzvlášť hojné v myelíne, struku, ktorý obklopuje a izoluje axóny niektorých neurónov.
Glykolipidy alebo neutrálne glykosfingolipidy (bez zaťaženia)
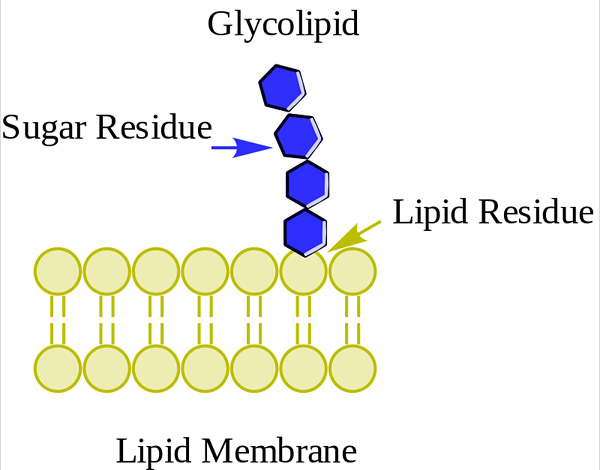 Glukolipid. Wpcrosson [CC BY-SA 4.0 (https: // creativeCommons.Org/licencie/By-SA/4.0)], z Wikimedia Commons
Glukolipid. Wpcrosson [CC BY-SA 4.0 (https: // creativeCommons.Org/licencie/By-SA/4.0)], z Wikimedia Commons Sú predovšetkým na vonkajšej tvári plazmovej membrány a majú jeden alebo viac cukrov ako polárna hlava priamo spojená s uhlíkom hydroxylom 1 ceramidovej časti. Nemajú fosfátové skupiny. Pretože pH 7 nemajú žiadne zaťaženie, nazývajú sa neutrálne glykolipidy.
Cerebrosidy majú jednu molekulu cukru spojenú s ceramidom. Tie, ktoré obsahujú galaktózu, sú v plazmatických membránach tkanivových buniek non -nerv. Globosidy sú goustoesfingolipidy s dvoma alebo viacerými cukrami, normálne d-glykózou, d-galaktózou alebo N-acetyl-d-galaktozamínom.
Gangliósidos alebo glykosfingolipidy
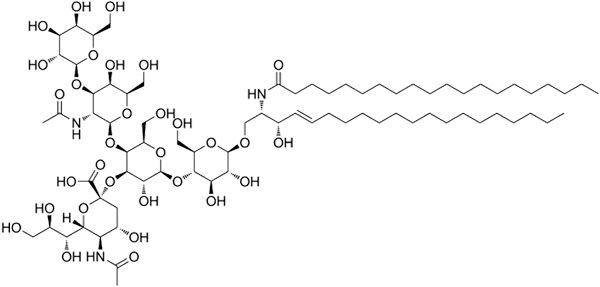 Štruktúra ganglosidov GM1
Štruktúra ganglosidov GM1 Toto sú najkomplexnejšie sfingolipidy. Majú oligosacharidy ako polárna skupina hlavy a jeden alebo viac terminálnych zvyškov kyseliny N-acetylmurálnej, tiež nazývané kyselina siral. Kyselina siatová prináša gangliám negatívne zaťaženie pH 7, ktoré ich odlišuje od neutrálnych glykosfingolipidov.
Nomenklatúra tohto druhu sfingolipidov závisí od množstva odpadu z kyseliny sinalovej prítomnej v oligosacharidovej časti polárnej hlavy.
Syntéza
Molekula s dlhým výskytom alebo sphiny Base je syntetizovaná v endoplazmatickom retikule (ER) a pridanie polárnej skupiny do hlavy týchto lipidov sa neskôr vyskytuje v komplexe Golgiho komplexu. U cicavcov sa v mitochondriách môže vyskytnúť aj niektoré syntéza sfingolipidov.
Po dokončení syntézy v komplexe Golgi sa sfingolipidy transportujú do iných bunkových kompartmentov mechanizmami sprostredkovanými vezikelmi.
Biosyntéza sfingolipidov pozostáva z troch základných udalostí: syntéza báz s dlhým reťazcom, biosyntéza ceramidov spojením mastnej kyseliny prostredníctvom väzby Amida a nakoniec tvorba komplexných sfingolipidov pomocou únie polárnych skupín v uhlíku 1 sfingoidovej bázy.
Okrem syntézy novo, Sfingolipidy sa môžu tiež tvoriť náhradou alebo recykláciou dlhých reťazcov a ceramidov, ktoré môžu kŕmiť sfingolipidový bazén.
Syntéza kostry ceramidu
Biosyntéza ceramidu, kostra sfingolipidov, začína dekarboxyleratívnou kondenzáciou molekuly palmitail-coA a L-serine. Reakcia je katalyzovaná heterodimérnym, pyridoxálnym fosfátom závislým a produktom je 3-Zo dihydroesfingosín Ze.
Môže vám slúžiť: Microbiota: Zloženie, funkcie a kde jeTento enzým je inhibovaný β-phaálnymi-L-alanínmi a L-Cicloserinas. V kvasinkách je kódovaný dvoma génmi, zatiaľ čo u cicavcov existujú tri gény pre tento enzým. Aktívne miesto je v cytoplazmatickej tvári endoplazmatického retikula.
Úloha tohto prvého enzýmu sa zachováva vo všetkých študovaných organizmoch. Existujú však určité rozdiely medzi taxónmi, ktoré sa týkajú subcelulárneho umiestnenia enzýmu: baktérie sú cytoplazmatické, kvasinky, rastliny a zvieratá sú v endoplazmatickom retikule.
3-zoesfinganín je následne znížený pomocou NADPH-dependent 3-Zo-ketosfinganín. Dihydroceramid syntázy (sfinganín N-acil prenosová) acetyl potom sfánanín na produkciu dihydroceramidu. Ceramid sa potom vytvorí pomocou dihydroceramidovej deaturázy/reduktázy, ktorá vkladá dvojitú väzbu previesť V pozícii 4-5.
U cicavcov je veľa syntasas pre ceramidové izoformy, z ktorých každá spája špecifický reťazec mastných kyselín s základňami dlhých reťazcov. Preto ceramidy syntézy a iné enzýmy, elonasy, poskytujú hlavný zdroj diverzity mastných kyselín v sfingolipidoch.
Metabolizmus
Degradáciu sfingolipidov vykonávajú glycohydroly a enzýmy sfingomyelinázy, ktoré sú zodpovedné za odstránenie modifikácií polárnych skupín. Na druhej strane, ceramidázy regenerujú základne dlhých reťazcov z ceramidov.
Gangliá sú degradované sadou lyzozomálnych enzýmov, ktoré katalyzujú krok za krokom cukrových jednotiek a nakoniec produkujú ceramid.
Ďalším spôsobom degradácie je internalizácia sfingolipidov v endocytických vezikulách, ktoré sa posielajú späť do plazmatickej membrány alebo transportované do lyzozómov, kde sú degradované špecifickou kyslou hydrolázou.
Nie všetky základne s dlhým reťazcom sa recyklujú, endoplazmatický zadržiavanie. Tento degradačný mechanizmus pozostáva z fosforylácie namiesto acilácie LCBS, čo vedie k signálnym molekulám, ktoré môžu byť rozpustné substráty pre enzým LASE.
Regulácia
Metabolizmus týchto lipidov je regulovaný na niekoľkých úrovniach, jednou z nich je enzýmy zodpovedné syntézou, ich post -translačných modifikácií a rovnakých mechanizmov rovnakých.
Niektoré regulačné mechanizmy sú špecifikácie buniek, buď na reguláciu času vývoja buniek, v ktorom sa vyrábajú, alebo v reakcii na konkrétne signály.
Odkazy
- Bartke, n., & Hannun, a. (2009). Bioaktívne spphingolipidy: metabolizmus a funkcia. Journal of Lipid Research, päťdesiat, 19.
- Breslow, D. Klimatizovať. (2013). Sfingolipidová homeostáza v endoplazmatickom retikule a ďalej. Perspektívy studeného jarného prístavu v biológii, 5(4), A013326.