

Reštrikčné enzýmy funkcie, typy a príklady
- 2050
- 513
- Tomáš Klapka
Ten obmedzené enzýmy Sú to endonukleázy používané určitými oblúkmi a baktériami na inhibíciu alebo „obmedzenie“ šírenia vírusov vo vnútri. Sú obzvlášť bežné v baktériách a sú súčasťou ich cudzieho systému obrany DNA známeho ako systém obmedzení/modifikácie.
Tieto enzýmy katalyzujú dvojitý pás DNA rezu na špecifických miestach, reprodukovateľné a bez použitia ďalšej energie. Väčšina z nich vyžaduje prítomnosť kofaktorov, ako je horčík alebo iné dvojmocné katióny, hoci niektoré tiež vyžadujú ATP alebo S-adenozyl metionín.
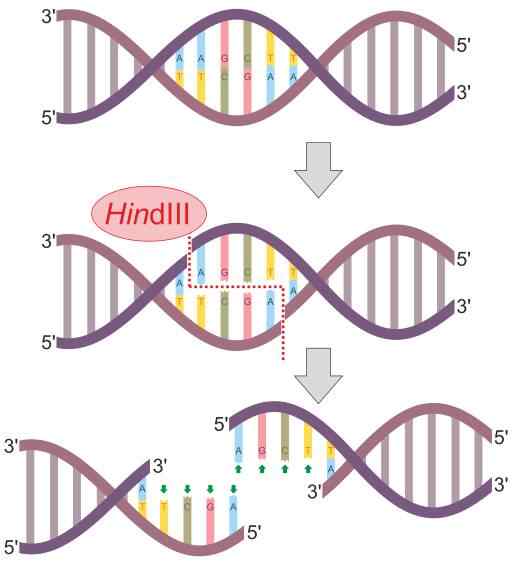
HINDII RESTRICTICKÁ ENZYME REAKCIA (Zdroj: Helixitta [CC BY-SA 4.0 (https: // creativeCommons.Org/licencie/By-SA/4.0)] Via Wikimedia Commons) Reštrikčné endonukleázy objavili v roku 1978 Daniel Nathans, Arber Werner a Hamilton Smith, ktorí získali Nobelovu cenu v medicíne za ich objav. Jeho meno vo všeobecnosti odchádza z organizmu, kde sú najprv pozorované.
Takéto enzýmy sa široko používajú pri vývoji metód klonovania DNA a inej molekulárnej biológie a stratégií genetického inžinierstva. Vďaka svojim charakteristikám rozpoznávania špecifických sekvencií a reznej kapacity sekvencií blízko k rozpoznávacím miestam z nich robia silné nástroje pri genetickom experimente.
Fragmenty generované reštrikčnými enzýmami, ktoré pôsobili na konkrétnu molekulu DNA, sa môžu použiť na obnovenie „mapy“ pôvodnej molekuly pomocou informácií na miestach, kde enzým preruší DNA.
Niektoré reštrikčné enzýmy môžu mať v DNA rovnaké rozpoznávacie miesto, ale nemusia ho nevyhnutne rezať rovnakým spôsobom. Existujú teda enzýmy, ktoré spôsobujú, že rezy zanechávajú Romos a enzýmy, ktoré rezajú opúšťajúce kohézne extrémy, ktoré majú rôzne aplikácie v molekulárnej biológii.
V súčasnosti sú komerčne dostupné stovky rôznych reštrikčných enzýmov, ktoré ponúkajú rôzne komerčné domy; Tieto enzýmy fungujú ako „prispôsobené“ molekulárne nožnice na rôzne účely.
[TOC]
Funkcia
Reštrikčné enzýmy plnia opačnú funkciu polymeráz, pretože sú hydrolyzované alebo lámajú esterovú väzbu v rámci fosfodiérovej väzby medzi susednými nukleotidmi v nukleotidovom reťazci.
V molekulárnej biológii a genetickom inžinierstve sú široko používané nástroje na konštrukciu expresných a klonovacích vektorov, ako aj na identifikáciu špecifických sekvencií. Sú tiež užitočné na konštrukciu rekombinantných genómov a majú veľký biotechnologický potenciál.
Nedávny pokrok v génovej terapii súčasne využíva reštrikčné enzýmy na zavedenie génov stanovených vo vektoroch, ktoré sú vozidlami na prepravu takýchto génov do živých buniek a že majú pravdepodobne schopnosť vložiť sa do bunkového genómu na vykonávanie trvalých zmien.
Mechanizmus akcie
Reštrikčné enzýmy môžu katalyzovať rezanie DNA s dvojitým pásmom, hoci niektoré sú schopné rozpoznať jednoduché sekvencie DNA pásma a dokonca aj RNA. Rez dochádza po rozpoznávaní sekvencií.
Môže vám slúžiť: glioxylátový cyklus: Charakteristiky, reakcie, regulácia, funkcieMechanizmus účinku pozostáva z hydrolýzy fosfodiérovej väzby medzi fosfátovou skupinou a deoxyribózou v kostre každej DNA DNA. Mnohé z enzýmov sú schopné rozrezať na rovnaké miesto, ktoré rozpoznávajú, zatiaľ čo iné sa rozrezajú medzi 5 a 9 páry báz pred alebo po tom istom.
Normálne tieto enzýmy rezané na 5 'konci fosfátovej skupiny, čo vedie k fragmentom DNA s 5' fosforyl koncom a koncom hydroxylu 3 '.
Pretože proteíny neprichádzajú do priameho kontaktu s miestom rozpoznávania DNA, musia byť translokované postupné časy, kým sa nedosiahne konkrétne miesto, možno prostredníctvom „posuvných“ mechanizmov na DNA Strand.
Počas enzymatického strihu je fosfodiérové spojenie každého zo vlákien DNA umiestnené v jednom z aktívnych miest reštrikčných enzýmov. Keď enzým opustí miesto rozpoznávania a rezania, robí tak prostredníctvom nešpecifických prechodných asociácií.
Chlapci
V súčasnosti je známych päť typov reštrikčných enzýmov. Ďalej stručný popis každého z nich:
Reštrikčné enzýmy typu I
Tieto enzýmy sú veľké pentamerické proteíny s tromi podjednotkami, jedným z obmedzení, metylácie a ďalšie pre rozpoznávanie sekvencií DNA. Tieto endonukleázy sú multifunkčné proteíny schopné katalyzujúcich reštrikčných a modifikačných reakcií, majú ATPASA a tiež aktivitu topoizomér DNA a tiež.
Enzýmy tohto typu boli prvými endonukleámi, ktoré sa objavili, boli po prvýkrát očistené v 60. rokoch a odvtedy boli študované s veľkou hĺbkou.
Enzýmy typu I sa široko nepoužívajú ako biotechnologický nástroj, pretože rezacím miestom môže byť variabilná vzdialenosť až do 1.000 párov báz týkajúcich sa lokality rozpoznávania, čo ich robí nespoľahlivými z hľadiska experimentálnej reprodukovateľnosti.
Reštrikčné enzýmy typu II
Sú to enzýmy zložené z homodimérov alebo tetramérov, ktoré rozrezávajú DNA na miesta definované medzi 4 a 8 bp dĺžky. Tieto rezacie miesta sú zvyčajne palindromické, to znamená, že rozpoznávajú sekvencie, ktoré sa čítajú rovnakým spôsobom v oboch smeroch.
Mnoho enzýmov reštrikčných enzýmov typu II v baktériách rezu DNA, keď rozpoznávajú svoj cudzí charakter, pretože nemá typické úpravy, ktoré by mala mať vlastná DNA.
Toto sú jednoduchšie reštrikčné enzýmy, pretože na rozpoznávanie a rezanie DNA sekvencií nevyžadujú iný kofaktor ako horčík (mg+).
Presnosť enzýmov reštrikčných enzýmov typu II pri rozpoznávaní a rezaní jednoduchých sekvencií v DNA v presných pozíciách z nich robí jeden z najpoužívanejších a najvýznamnejších vo väčšine vetiev molekulárnej biológie.
Môže vám slúžiť: vzájomný vzťah: Charakteristiky, typy, príkladyV skupine enzýmov reštrikcie typu II sú viacnásobné podtriedy klasifikované podľa určitých vlastností, ktoré sú pre každú jedinečnú. Klasifikácia týchto enzýmov sa vykonáva pridaním písmen abecedy od A do Z podľa názvu enzýmu.
Niektoré z najznámejších podtriedy ich užitočnosti sú:
Podtrieda IIA
Sú to rôzne podjednotky Dímeros. Rozpoznávajú asymetrické sekvencie a používajú sa ako ideálne prekurzory na generovanie rezných enzýmov.
Podtrieda IIB
Skladajú sa z jedného ďalšieho diméru a odrezajú DNA na oboch stranách rozpoznávacej sekvencie. Neskôr z miesta rozpoznávania odrezali obe pramene DNA v intervale párov báz neskôr.
IIC podtrieda
Enzýmy tohto typu sú polypeptidy s funkciami delenia a modifikácie prameňov DNA. Tieto enzýmy rezajú obidve pramene asymetricky.
Podtrieda IIE
Enzýmy tejto podtriedy sú najpoužívanejšie v genetickom inžinierstve. Majú katalytické miesto a vo všeobecnosti vyžadujú efektor Alestro. Tieto enzýmy musia interagovať s dvoma kópiami ich rozpoznávacej sekvencie, aby sa dosiahlo efektívne rezanie. V rámci tejto podtriedy sú enzýmy ekorii a ekori.
Reštrikčné enzýmy typu III
Reštrikčné endonukleázy typu III sú zložené iba z dvoch podjednotiek, jedna je zodpovedná za rozpoznávanie a modifikáciu DNA, zatiaľ čo druhá je zodpovedná za reznosť sekvencie.
Tieto enzýmy vyžadujú na svoju prevádzku dva kofaktory: ATP a horčík. Reštrikčné enzýmy tohto typu majú dve asymetrické rozpoznávacie miesta, premiestňujte DNA spôsobom závislým od ATP a znižuje ju medzi 20 až 30 bp susediacimi s miestom rozpoznávania.
Reštrikčné enzýmy typu IV
Enzýmy typu IV sa dajú ľahko identifikovať, pretože DNA rezajú pomocou metylačných značiek, tvoria sa z niekoľkých rôznych podjednotiek, ktoré sú zodpovedné za rozpoznávanie a rezanie sekvencie DNA. Tieto enzýmy používajú gtp kofaktory a dvojité horčík.
Medzi špecifické rezacie miesta patrí nukleotidové reťazce s metylovanými alebo hydroxymetylovanými cytozínovými zvyškami v jednom alebo oboch vláknach nukleových kyselín.
Enzýmy obmedzenia typu V
Táto klasifikačná skupina zoskupuje enzýmy typu Crossper-Cas, ktoré identifikujú a znižujú špecifické DNA sekvencie inváznych organizmov. Enzýmy CAS používajú sprievodcu syntetizovaným RNA prameňom pre Crisper na rozpoznávanie a útočenie na inváziu organizmov.
Enzýmy klasifikované ako typ V sú polypeptidy štruktúrované pomocou enzýmov typu I, II a II. Môžu rezať sekcie DNA takmer akéhokoľvek organizmu a s veľkou škálou dĺžky. Vďaka svojej flexibilite a ľahkom zamestnaní sú tieto enzýmy jedným z najpoužívanejších nástrojov v genetickom inžinierstve v súčasnosti spolu s enzýmami typu II.
Môže vám slúžiť: propionibacteriumPríklady
Na detekciu polymorfizmov DNA sa použili reštrikčné enzýmy, najmä v populačných genetických štúdiách a evolučných štúdiách využívajúcich mitochondriálnu DNA, aby sa získali informácie o miere substitúcie nukleotidov.
V súčasnosti majú vektory používané na transformáciu baktérií s rôznymi účelmi, kde sa nachádzajú miesta rozpoznávania viacerých reštrikčných enzýmov.
Medzi týmito enzýmami najobľúbenejšie sú Ecori, II, III, IV a V, získané a opísané prvýkrát z A. coli; Hindii z H. influinzae a Bamhi of B. Amyloliquefaciens.
Odkazy
- Bikle, t. Do., & Kruger, D. H. (1993). Biológia obmedzenia DNA. Mikrobiologické recenzie, 57(2), 434-450.
- Boyval, P., Moineau, s., Romero, D. Do., & Horvath, P. (2007). CRISPR poskytuje nadobudnutie proti vírusom v prokaryotoch. Veda, 315(Marec), 1709-1713.
- Goodsell, D. (2002). Molekulárna perspektíva: Reštrikčné endonukleázy. Kmeňové bunky Základy rakovinovej medicíny, dvadsať, 190-191.
- Halford, s. A. (2001). Poskakovanie, skákanie a opakovanie reštrikčnými enzýmami. Transakcie biochemickej spoločnosti, 29, 363-373.
- Jeltsch, a. (2003). Údržba druhovej identity a kontrola špekulácie baktérií: nová funkcia pre systémy obmedzenia/modifikácie? Gén, 317, 13-16.
- Krebs, J., Goldstein, e., & Kilpatrick, s. (2018). Lewinove gény XII (12 ed.). Burlington, Massachusetts: Jones & Bartlett Learning.
- Li, a., Chlieb, s., Zhang a., Ren, m., Feng, m., Peng, n.,... ona, Q. (2015). Využívanie systémov typu I a typu III CRISPR-CAS pre úpravy genómu. Výskum nukleových kyselín, 1-12.
- Loenen, W. Do. M., Dryden, D. Tón. F., Raleight, e. Do., & Wilson, G. G. (2013). Enzýmy reštrikcie typu I a ich príbuzní. Výskum nukleových kyselín, 1-25.
- Nathans, D., & Smith, h. Ani. (1975). Reštrikčné endonukleázy v analýze a reštrukturalizácii molekúl DNA. Anu. Otáčať sa. Biochem., 273-293.
- Nei, m., & Tajima, f. (1981). DNA polymorfizmus detegovateľný reštrikčnými endonukleázami. Genetika, 145-163.
- Pingoud, a., Fuxreiter, m., Pingoud, v., & Wende, w. (2005). Bunkové a molekulárne vedy o životnosti typu II Reštrikčné endonukleázy: Štruktúra a mechanizmus. CMLS bunkové a molekulárne vedy, 62, 685-707.
- Roberts, r. (2005). Ako sa reštrikčné enzýmy stali pracovnými koncami molekulárnej biológie. Pnas, 102(17), 5905-5908.
- Roberts, r. J., & Murray, K. (1976). Reštrikčné endonukleázy. Kritické prehľady v biochémii, (November), 123-164.
- Stoddard, B. L. (2005). Štruktúra a funkcia endonukleázy navádzania. Štvrťročné recenzie biofyziky, 1-47.
- Tock, m. R., & Dryden, D. Tón. F. (2005). Biológia obmedzenia a protivládania. Súčasný názor v mikrobiológii, 8, 466-472. https: // doi.org/10.1016/j.Môj b.2005.06.003
- Wilson, G. G., & Murray, n. A. (1991). Systémy obmedzenia a modifikácie. Anu. Otáčať sa. Genet., 25, 585-627.
- Wu, z., & Mou, k. (2016). Genomické poznatky o virulencii a populačnej genetike Campylobacter. Infekcia. Disk. Preložiť. Prezerať., 2(3), 109-19.
- Yuan, r. (1981). Štruktúra a mechanizmus multifunkčných reštrikčných endonukleánov. Anu. Otáčať sa. Biochem., päťdesiat, 285-315.
- « Generovanie charakteristík, autorov a diel z roku 1914
- Vlastnosti a príklady vzájomne sa vylučujú udalosti »