

Štruktúra mastných kyselín, typy, funkcie, biosyntéza
- 4406
- 41
- Alan Milota
Ten mastné kyseliny Sú to organické makromolekuly odvodené od uhľovodíkov, ktoré sa skladajú z dlhých reťazcov atómov uhlíka a vodíka, ktoré sú hydrofóbne (sú rozpustné v tuku) a sú štruktúrnym základom tukov a lipidov.
Sú to veľmi rozmanité molekuly, ktoré sa navzájom odlišujú dĺžkou ich uhľovodíkových reťazcov a prítomnosťou, číslom, polohou a/alebo konfiguráciou ich dvojitých väzieb.
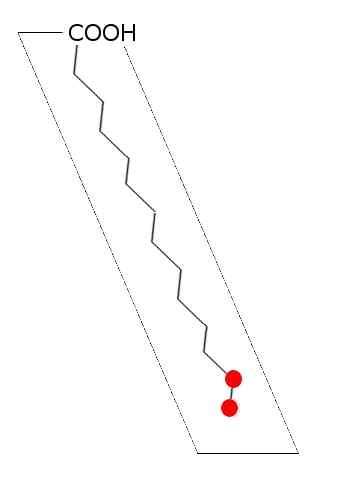
 Všeobecná schéma nasýtenej mastnej kyseliny (zdroj: Laghi.L [CC BY-SA 3.0 (http: // creativeCommons.Org/licencie/By-SA/3.0/)] Via Wikimedia Commons)
Všeobecná schéma nasýtenej mastnej kyseliny (zdroj: Laghi.L [CC BY-SA 3.0 (http: // creativeCommons.Org/licencie/By-SA/3.0/)] Via Wikimedia Commons) U lipidov zvierat, rastlín, húb a mikroorganizmov, ako sú baktérie a kvasinky, bolo opísaných viac ako 100 druhov rôznych mastných kyselín a u väčšiny živých bytostí sa považujú za špecifické druhy a tkanivo.
Oleje a tuky, ktoré človek konzumuje denne, sú pôvodom zvierat alebo rastlín, väčšinou sa skladajú z mastných kyselín.
[TOC]
Zavedenie
 Maslo, okrem iného, sú v podstate zložené z mastných kyselín (zdroj: Africa Studio, Via Pixabay.com)
Maslo, okrem iného, sú v podstate zložené z mastných kyselín (zdroj: Africa Studio, Via Pixabay.com) Molekuly tukových kyselín vykonávajú dôležité funkcie na bunkovej úrovni, čo z nich robí základné zložky a keďže niektoré z nich nemôžu byť syntetizované zvieratami, musia ich získať z diéty.
Mastné kyseliny sú zriedkavé ako voľné druhy v bunkovom cytosole, takže sú vo všeobecnosti súčasťou iných molekulárnych konjugátov, ako napríklad:
- Lipidy, v biologických membránach.
- Triglyceridy alebo estery mastných kyselín, ktoré slúžia ako rezerva v rastlinách a zvieratách.
- Vosky, ktoré sú tuhé estery mastných kyselín a alkoholov s dlhým reťazcom.
- Ďalšie podobné látky.
U zvierat sa mastné kyseliny ukladajú v cytoplazme buniek ako malé kvapky tuku zloženého z komplexu nazývaného triacylglycerol, ktorý nie je ničím iným ako molekula glycerolu, ku ktorej sa spojila, v každom zo svojich atómov uhlíka, reťazca mastných kyselín Podľa väzieb typu esteru.
Zatiaľ čo baktérie majú krátke a zvyčajne mononenasýtené mastné kyseliny, v prírode je bežné.
Štruktúra
Mastné kyseliny sú amfipatické molekuly, to znamená, že majú dve chemicky definované oblasti: hydrofilná polárna oblasť a hydrofóbna apolárna oblasť.
Hydrofóbna oblasť sa skladá z dlhého uhľovodíkového reťazca, ktorý z chemického hľadiska nie je príliš reaktívny. Na druhej strane hydrofilná oblasť sa skladá z terminálu karboxylovej skupiny (-COH), ktorá sa správa ako kyselina.
Táto terminálna karboxylová skupina alebo karboxylová kyselina je ionizovaná v roztoku, je vysoko reaktívna (chemicky povedané) a je veľmi hydrofilná, takže predstavuje kovalentné spojenie medzi mastnými kyselinami a inými molekulami.
Dĺžka uhľovodíkových reťazcov mastných kyselín má zvyčajne počet atómov uhlíka, a to úzko súvisí s biosyntetickým procesom, ktorým sa vyrábajú, pretože ich rast sa vyskytuje z párov uhlíkov.
Najbežnejšie mastné kyseliny majú reťazce medzi 16 a 18 atómami uhlíka a zvieratám, tieto reťazce nie sú rozvetvené.
Klasifikácia
Mastné kyseliny sa klasifikujú do dvoch veľkých skupín podľa povahy spojení, ktoré ich tvoria, to znamená podľa prítomnosti jednoduchých väzieb alebo dvojitých väzieb medzi atómami uhlíka ich uhľovodíkových reťazcov.
Preto sú nasýtené a nenasýtené mastné kyseliny.
- Nasýtené mastné kyseliny majú iba jednoduché uhlíkové väzby a všetky ich atómy uhlíka sú „nasýtené“ alebo spojené s molekulami vodíka.
- Nenasýtené mastné kyseliny majú jednu alebo viac dvojitých uhlíkových väzieb a nie všetky sú pripojené k atómu vodíka.
Nenasýtené mastné kyseliny sú tiež rozdelené podľa počtu nenasýtení (dvojité väzby) v mononenasýtených, tých s jednou dvojitou väzbou a polynenasýtené, ktoré majú viac ako jednu.
Nasýtené mastné kyseliny
Zvyčajne majú medzi 4 a 26 atómami uhlíka spojené jednoduchými väzbami. Jeho bod topenia je priamo úmerný dĺžke reťazca, to znamená jeho molekulovej hmotnosti.
Mastné kyseliny, ktoré majú 4 až 8 uhlíkov, sú kvapalné pri 25 ° C a sú to tie, ktoré tvoria jedlé oleje, zatiaľ čo tie, ktoré majú viac ako 10 atómov uhlíka, sú pevné.
Medzi najbežnejšie patrí kyselina laurová, ktorá je hojná v semenách dlane a kokosu; Kyselina palmitová, ktorá sa nachádza v dlani, kakao a masle ošípaných a kyselinou stearová, ktorá je v kakao a hydrogenovaných olejoch.
Sú to mastné kyseliny s oveľa väčšou stabilitou ako nenasýtené mastné kyseliny, najmä vzhľadom na oxidáciu, prinajmenšom vo fyziologických podmienkach.
Vďaka skutočnosti, že jednoduché väzby uhlíka - uhlíka sa môžu striedať zadarmo.
Nenasýtené mastné kyseliny
Tieto mastné kyseliny sú vysoko reaktívne a náchylné k saturácii a oxidácii. Sú bežné v rastlinách a morských organizmoch. Tí, ktorí majú jediné dvojité puto, sú známe ako mononenasýtené alebo monoenoické, zatiaľ čo tí, ktorí majú viac ako dva, sú známi ako polyenika alebo polynasýtení.
Môže vám slúžiť: DNA polymerázaPrítomnosť dvojitých väzieb je bežná medzi atómami uhlíka medzi polohami 9 a 10, to však neznamená, že monouinsturované mastné kyseliny sa nenachádzajú s nenasýtením v inej polohe.
Na rozdiel od nasýtených, nenasýtených mastných kyselín nie sú uvedené nie z terminálnej karboxylovej skupiny, ale podľa polohy prvej dvojitej väzby C - C. Tieto sú teda rozdelené do dvoch skupín, omega-6 alebo co6 a omega-3 alebo co3.
Kyseliny omega-6 majú prvú dvojitú väzbu v uhlíkovom čísle 6 a kyseliny omega-3 ho majú v uhlíkovom čísle 3. Henovina Ω je daná dvojitou väzbou najbližšie k konečnej metylovej skupine.
Dvojité odkazy nájdete aj v dvoch geometrických konfiguráciách známych ako “Cis " a „Trans“.
Väčšina prírodných nenasýtených mastných kyselín má konfiguráciu „CIS”A dvojité väzby mastných kyselín prítomných v komerčných tukoch (hydrogenované) sa nachádzajú v „Trans“.
V polynenasýtených mastných kyselinách sú dve dvojité väzby obvykle od seba oddelené aspoň metylovou skupinou, to znamená atóm uhlíka pripojený k dvom atómom vodíka.
Funkcia
Mastné kyseliny majú viac funkcií v živých organizmoch a, ako je uvedené vyššie, jednou z ich základných funkcií ako podstatnej súčasti lipidov, ktoré sú hlavnými zložkami biologických membrán a jednou z troch najhojnejších biomolekúl v organizmoch proteíny a uhľohydráty.
Sú tiež vynikajúcimi energetickými substrátmi, vďaka ktorým sa získavajú veľké množstvo energie vo forme ATP a iných sprostredkovateľských metabolitov.
Vzhľadom na skutočnosť, že zvieratá napríklad nie sú schopné ukladať uhľohydráty, mastné kyseliny predstavujú hlavný zdroj skladovania energie, ktorý pochádza z oxidácie nadbytočných cukrov.
Mastné mastné kyseliny s krátkym reťazcom v hrubom čreve sa podieľajú na stimulácii absorpcie vody a sodíka, chloridu a iónov hydrogenuhličitanu; Okrem toho majú funkcie pri výrobe hlienu, v proliferácii kolonocytov (bunky hrubého čreva) atď.
Nenasýtené mastné kyseliny sú obzvlášť hojné v jedlých olejoch rastlinného pôvodu, ktoré sú dôležité v strave všetkých ľudí.
 Oleje, ktoré denne konzumujeme, sú mastné kyseliny (zdroj: Stevepb, cez Pixabay.com)
Oleje, ktoré denne konzumujeme, sú mastné kyseliny (zdroj: Stevepb, cez Pixabay.com) Iní sa zúčastňujú ako ligandy niektorých proteínov s enzymatickými aktivitami, takže sú dôležité, pokiaľ ide o ich účinky na energetický metabolizmus buniek, kde sa nachádzajú.
Biosyntéza
Degradácia mastných kyselín je známa ako β-oxidácia a vyskytuje sa v mitochondriách eukaryotických buniek. Naopak biosyntéza sa vyskytuje v cytosóle živočíšnych buniek a v chloroplastoch (fotosyntetické organely) rastlinných buniek.
Je to proces acetyl-CoA, Malonyl-CoA a NADPH, vyskytuje sa vo všetkých živých organizmoch a u „vynikajúcich“ zvierat, ako sú cicavce. Napríklad je veľmi dôležitý v pečeni a tukových tkanivách, ako aj v prsných žľazach.
NADPH používaný na túto cestu je hlavne produktom oxidačných reakcií závislých od NADP, zatiaľ čo acetyl-CoA môže pochádzať z rôznych zdrojov, napríklad z oxidačnej dekarboxylácie pyruvátu, Krebs a p-β- oxidácia mastných kyselín.
Trasa biosyntézy, podobne ako β-oxidácia, je vysoko regulovaná vo všetkých bunkách astherskými efektormi a kovalentnými modifikáciami enzýmov, ktoré sa zúčastňujú na regulácii.
-Syntéza Malonyl-CoA
Trasa začína tvorbou metabolického sprostredkovateľa známeho ako Malonyl-CoA z molekuly acetyl-CoA a je katalyzovaná multifunkčným enzýmom nazývaným acetyl-CoA carboxilasa.
Táto reakcia je reakciou na pridanie karboxylovej molekuly (-COH, karboxylácia) v závislosti od biotínu a vyskytuje sa v dvoch krokoch:
- Po prvé, prenos, závislý od ATP, karboxylového odvodeného z bikarbonátu (HCO3-) do molekuly biotínu, ktorá sa nachádza ako protetická (neproteín) skupina spojená s acetyl-COA karboxylázou.
- Následne sa CO2 prenesie na acetyl-CoA a vyrába sa Malonyl-CoA.
-Reakcie trasy
U zvierat sa k tvorbe uhľohydrátových reťazcov mastných kyselín dochádza ďalej prostredníctvom sekvenčných kondenzačných reakcií katalyzovaných multikritickým a multifunkčným enzýmom, ktorý je známy ako syntázia mastných kyselín.
Tento enzým katalyzuje kondenzáciu acetyl-CoA jednotky a molekúl s viacerými COA, ktoré sa vyrábajú z reakcie karboxylázy acetyl-CoA, proces, počas ktorého sa molekula CO2 uvoľňuje pre každý malonyl-CoA, ktorý pridáva.
Pestovanie mastných kyselín sú sterifikované na bielkoviny nazývané „proteín s acylovým nosičom“ alebo ACP, ktorý tvorí tioé s acylovými skupinami. V A. coli Tento proteín je 10 kDa polypeptid, ale u zvierat je súčasťou syntázy mastných kyselín.
Môže vám slúžiť: NoxaRuptúra týchto dlhopisov Tioéster uvoľňuje veľké množstvo energie, čo je možné, termodynamicky povedané, výskyt kondenzačných krokov na biosintetickej trase.
Syntáza komplexu mastných kyselín
V baktériách aktivita syntázovej kyseliny v skutočnosti zodpovedá šiestim nezávislým enzýmom, ktoré používajú acetyl-CoA a malonyl-CoA na vytvorenie mastných kyselín a s ktorými je spojených šesť rôznych enzymatických aktivít.
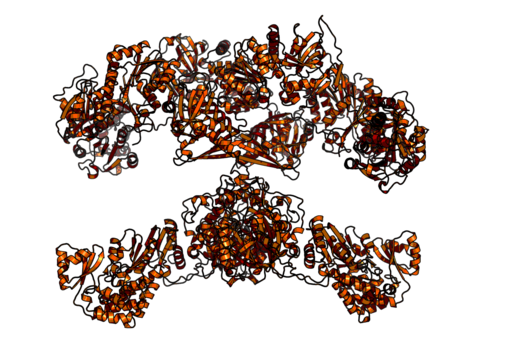 Homodimérna a multifunkčná komplexná mastná kyselina syntáza zvierat (zdroj: Boehringer Ingelheim [CC BY-SA 4.0 (https: // creativeCommons.Org/licencie/By-SA/4.0)] Via Wikimedia Commons)
Homodimérna a multifunkčná komplexná mastná kyselina syntáza zvierat (zdroj: Boehringer Ingelheim [CC BY-SA 4.0 (https: // creativeCommons.Org/licencie/By-SA/4.0)] Via Wikimedia Commons) Na druhej strane u cicavcov je syntázia mastných kyselín multifunkčným homodimitovým enzymatickým komplexom s molekulovou hmotnosťou približne 500 kDa, ktorý má šesť rôznych katalytických aktivít a s ktorým je kyslý nosný proteín spojený.
Krok 1: Reakcia Prowter
Tiolové skupiny v cysteínovom odpade zodpovedným za spojenie metabolických sprostredkovateľov do enzýmu ACP sa musia načítať pred začiatkom syntézy, s potrebnými kyslými skupinami.
Na tento účel sa acetyl-CoA acetyl skupina prenáša do skupiny TIOL (-SH) jedného z cysteínových zvyškov podjednotky ACP syntázy mastných kyselín. Táto reakcia je katalyzovaná podjednotkou ACP-acil-transferáza.
Acetylová skupina sa potom prenesie z ACP do iného cysteínového zvyšku v katalytickom mieste inej enzymatickej podjednotky komplexu, ktorý je známy ako β-cethoacyl-ACP-Sinta. Enzymatický komplex je teda „jačmeň“ na začatie syntézy.
Krok 2: Prenos jednotiek Malonyl-CoA
Malonyl-CoA, ktorý vyrába acetyl-CoA carboxilasa, sa prenesie do skupiny TIOL v ACP a počas tejto reakcie sa časť COA stratí. Reakcia je katalyzovaná podjednotkou Malonil-ACP-transferázy komplexu mastných kyselín, ktorá potom produkuje malonil-ACP.
Počas tohto procesu je malonická skupina spojená s ACP a β-kotoacil-ACP-SITA prostredníctvom esterového spojenia a iného sulfidlu.
Krok 3: Kondenzácia
Enzým β-zoacil-ACP-Suntázy katalyzuje prenos acetylovej skupiny, ktorá sa k nemu pripojila pri prechode „jačmeňa“ smerom k uhlíkovi 2 skupiny Malonilo, ktorý bol v predchádzajúcom kroku prenesený na AKT do ACP.
Počas tejto reakcie sa uvoľní molekula CO2, ktorá pochádza z Malonyla, čo zodpovedá CO2 prispel bikarbonátom v karboxylačnej reakcii karboxylázy acetyl-CoA. Potom sa vyskytuje acetoacetyl-ACP.
Krok 4: Redukcia
Podjednotka p-zoacil-Acp-reduktázy katalyzuje redukciu Aceoacetyl-ACP závislé od NADPH.
Krok 5: Dehydratácia
Tento krok tvorí trans-a, β-acyl-ACP alebo ∆2-insaturado-acil-ACP (cratonil-ACP), produkt dehydratácie D-β-hydroxybutiril-ACP pôsobením nahnevanej podjednotky ACP -Hydratasa.
Neskôr sa cratonil-ACP redukuje na butiril-ACP reakciou v závislosti od NADPH katalyzovanej podjednotkou nahnevanej ac-reduktázy. S touto reakciou je prvý zo siedmich cyklov potrebných na výrobu ACP palmitail, ktorý je predchodcom takmer všetkých mastných kyselín.
Ako nasledujú následné kondenzačné reakcie?
Skupina Butirilo sa prenáša z ACP do skupiny TIOL cysteínového zvyšku v β-cethoacil-ACP-SITA, takže ACP je schopný prijať ďalšiu malonickú skupinu z Malonyl-CoA.
Týmto spôsobom je reakcia, ktorá sa stane, kondenzácia malonil-ACP pomocou buturil-β-kotoacil-ACP-sinta, ktorá vedie k vzostupu β-zotoohexanoil-ACP + CO2.
Palmitail-ACP, ktorý vzniká z nasledujúcich krokov (po pridaní 5 ďalších malonických jednotiek), sa môže uvoľniť ako kyselina palmitová vďaka aktivite enzýmu tioesterázy, môže sa preniesť do COA alebo začleniť do kyseliny fosfatidovej kyseliny Syntéza cesty fosfolipidov a triacylglyceridov.
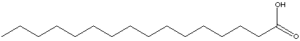 Štruktúra kyseliny palmitovej (zdroj: Andel, cez Wikimedia Commons)
Štruktúra kyseliny palmitovej (zdroj: Andel, cez Wikimedia Commons) Syntáza mastných kyselín väčšiny organizmov sa zastaví v syntéze ACP palmitail-ACP, vzhľadom na to, že katalytické miesto podjednotky β-cethoacyl-ACP-syntázy má konfiguráciu, v ktorej sa môžu prispôsobiť iba mastné kyseliny tejto dĺžky.
Ako sú mastné kyseliny nepárnych počtu atómov uhlíka?
Sú relatívne bežné v morských organizmoch a sú syntetizované aj komplexnou syntázou mastných kyselín. Reakcia „jačmeňa“ sa však vyskytuje s dlhšou molekulou, propionyl-ACP, troch atómov uhlíka.
Kde a ako sú mastné kyseliny dlhších reťazcov?
Kyselina palmitová, ako bolo komentované, slúži ako predchodca mnohých nasýtených a nenasýtených mastných kyselín s dlhšími reťazcami. Proces „predĺženia“ mastných kyselín sa vyskytuje v mitochondriách, zatiaľ čo zavedenie nenasýtenia sa vyskytuje v podstate v endoplazmatickom retikule.
Mnoho organizmov premení svoje nasýtené mastné kyseliny na nenasýtené ako prispôsobenie sa nízkym teplotám prostredia, pretože to umožňuje udržiavať fúzny bod lipidov pod okolitou teplotou.
Vlastnosti mastných kyselín
Mnoho vlastností mastných kyselín závisí od dĺžky ich reťazca a prítomnosti a počtu nenasýtení:
Môže vám slúžiť: teória abiotickej syntézy: hlavné charakteristiky- Nenasýtené mastné kyseliny majú nižšie body topenia ako nasýtené mastné kyseliny s rovnakou dĺžkou.
- Dĺžka mastných kyselín (počet atómov uhlíka) je nepriamo úmerná plynulosti alebo flexibilite molekuly, to znamená, že najviac „krátke“ molekuly sú plynulejšie a naopak.
Všeobecne platí, že mastné látky tekutí.
Rastliny majú hojné množstvo nenasýtených mastných kyselín, ako aj zvieratá, ktoré žijú pri veľmi nízkych teplotách, pretože tieto, ako zložky lipidov prítomných v bunkových membránach, im v týchto podmienkach poskytujú väčšiu plynulosť.
Vo fyziologických podmienkach prítomnosť dvojitej väzby v uhľovodíkovom reťazci mastného.
Prítomnosť dvojitých väzieb v mastných kyselinách spojených s molekulami lipidov má priame účinky na stupeň „balenia“, ktoré môžu mať v membránach, ku ktorým patria, a s ním tiež majú účinky na membránové proteíny.
 Príklad tvorby myla mastných kyselín s karboxylovými skupinami vystavenými do vodného prostredia (zdroj: Benutzer: Anderl [CC BY-SA 3.0 (http: // creativeCommons.Org/licencie/By-SA/3.0/)] Via Wikimedia Commons)
Príklad tvorby myla mastných kyselín s karboxylovými skupinami vystavenými do vodného prostredia (zdroj: Benutzer: Anderl [CC BY-SA 3.0 (http: // creativeCommons.Org/licencie/By-SA/3.0/)] Via Wikimedia Commons) Rozpustnosť mastných kyselín klesá so zvyšovaním dĺžky jeho reťazca, takže sú nepriamo úmerné. Vo vodných a lipidových zmesiach sú mastné kyseliny spojené v štruktúrach známych ako micely.
Micella je štruktúra, v ktorej sú alifatické reťazce mastných kyselín „uzavreté“, čím „vylučuje“ všetky molekuly vody a na ktorej povrch.
Menovanie
Nomenklatúra mastných kyselín môže byť trochu zložitá, najmä ak sa týka bežných mien, ktoré dostávajú, ktoré často súvisia s niektorými fyzikálno -chemickými vlastnosťami, s miestom, kde sú alebo iné vlastnosti.
Mnoho autorov sa domnieva, že keďže vďaka terminálovej karboxylovej skupine sú tieto molekuly ionizované na fyziologické pH, mali by sa ich označovať ako „karboxilates“ pomocou ukončenia “Ato ".
Podľa systému IUPAC je vymenovanie atómov uhlíka tukového. Reťazový terminálny metyl obsahuje atóm uhlíka Ω.
Všeobecne platí, že v systematickej nomenklatúre dostávajú názov „rodičovského“ uhľovodíka (uhľovodík s rovnakým počtom atómov uhlíka) a ich ukončenie sa nahradí “ani„ „Oico“, Ak ide o nenasýtenú mastnú kyselinu, dokončenie sa pridá „Hmla“.
Zoberme si napríklad prípad mastnej kyseliny 18 atómov uhlíka (C18):
- Pretože uhľovodík s rovnakým množstvom atómov uhlíka je známy ako Octadecano, nasýtená kyselina sa nazýva “Kyselina oktadekanová„Ó dobre“Oktadecanoato“A jej spoločným názvom je kyselina stearová.
- Ak máte v jej štruktúre dvojitú väzbu medzi niektorými pármi atómov uhlíka, je známa ako “Kyselina oktadenoová„
- Ak máte dve dvojité c res c - c, potom sa volá „Kyselina oktadekadienoová“ A ak máte tri “Kyselina oktadekatrináová„.
Ak chcete zhrnúť nomenklatúru, potom použite 18: 0 pre 18 -karbonové mastné kyseliny a bez dvojitej väzby (nasýtená) a v závislosti od stupňa nenasýtenia, namiesto nuly je napísaná 18: 1 pre molekulu s molekulou s nenasýtením, 18: 2 pre jednu s dvoma nenasýtením a tak ďalej.
Ak chcete špecifikovať medzi tým, ktoré atómy uhlíka sú dvojité väzby v nenasýtených mastných kyselinách, symbol ∆ sa používa s numerickým prieskumom, ktorý naznačuje miesto nenasýtenia a predpony „CIS„Buď „Trans“, v závislosti od konfigurácie tohto.
Odkazy
- Badui, s. (2006). Chémia potravín. (A. Quintanar, ed.) (4. vydanie.). Mexiko d.F.: Pearson Education.
- Garrett, R., & Grisham, C. (2010). Biochémia (4. vydanie.). Boston, USA: Brooks/Cole. Učenie sa.
- Mathews, C., Van Holde, K., & Ahern, K. (2000). Biochémia (3. vydanie.). San Francisco, Kalifornia: Pearson.
- Murray, r., Bender, D., Botham, K., Kennelly, P., Rodwell, V., & Weil, P. (2009). Harperova ilustrovaná biochémia (28. ED.). McGraw-Hill Medical.
- Nelson, D. L., & Cox, M. M. (2009). Lehninger princípy biochémie. Vydanie omega (5. vydanie.).
- Rawn, J. D. (1998). Biochémia. Burlington, Massachusetts: Vydavatelia Neil Patterson.
- Tvrzicka, e., Kremmyda, l., Stankovia, b., & Zak,. (2011). Mastné kyseliny ako biokompounds: ich úloha v ľudskom metabolizme, zdraví a chorobe- prehľad. Časť 1: Klasifikácia, strava a biologické funkcie. Biomed Pap Med Fac Univ Palacky Olomouc Česka repub, 155(2), 117-130.
- « Štruktúra glykoproteínov, funkcie, triedy a príklady
- Vplyv príslušnosti k digitálnemu občianstvu »