

Charakteristiky aktínu, štruktúra, vlákna, funkcia
- 1971
- 317
- Adrián Fajnor
Ten Aktín Je to cytosolický proteín, ktorý tvorí mikrofilamenty. V eukaryotoch je aktín jedným z najhojnejších proteínov. Napríklad predstavuje 10% hmotnosť celkového proteínu vo svalových bunkách; a medzi 1 a 5% proteínu v bunkách ne -muscle.
Tento proteín, vedľa stredných vlákien a mikrotubulov, tvorí cytoskelet, ktorého hlavnou funkciou je mobilita bunky, udržiavanie bunkovej formy, bunkové delenie a pohyb organel v rastlinách, húb a zvieratách.
Zdroj: Sarcomere.SVG: David Richfield (užívateľ Slashme) Derivátová práca: Retama [CC BY-SA 3.0 (http: // creativeCommons.Org/licencie/By-SA/3.0/]] Izoformy aktínového cytoskelet.
Z evolučného hľadiska je aktín vysoko konzervovaný proteín. Homológia sekvencie je asi 90% u rôznych druhov. V jednobunkových organizmoch jeden gén kóduje aktín Isaform. Zatiaľ čo v mnohobunkových organizmoch, rôzne gény kódujú viacero aktínových Isaformy.
Actín spolu s myozínom bol rozhodujúcimi štruktúrami v evolučnej budúcnosti eukaryotických organizmov a ich diverzifikácie, pretože umožňovali pohyb v neprítomnosti iných štruktúr, ako sú bičíky a cilia.
[TOC]
Štruktúra: Actínové vlákna
Aktín je jediný globulárny proteín polypeptidového reťazca. Vo svale má aktín molekulovú hmotu približne 42 kDa.
Tento proteín má dve domény. Každá z nich má dve subdomény a rozštiepenie medzi doménami. ATP-MG+2 pripojí sa na spodok rozštepu. Terminály amino a karboxilu sa stretávajú v subdoméne 1.
Actina G a Actina f
Existujú dve hlavné aktínové formy: Actin monomér, nazývaný Actina G; a vláknitý polymér, ktorý sa skladá z aktínov g monomérov, nazývaný Actina f. Aktínové vlákna, pozorované elektronickou mikroskopiou, majú úzke a široké oblasti, respektíve priemer 7 nm a 9 nm.
Počas celého vlákna tvorí Actin Monomers dvojitú špirálu. Jednotka, ktorá sa opakuje v celom vlákne, sa skladá z 13 vrtule a 28 aktínových monomérov a má vzdialenosť 72 nm.
Actínové vlákno má dva ciele. Jeden tvorí rozštep, ktorý spája ATP-MG+2, ktorý sa nachádza rovnakým smerom vo všetkých aktínskych monoméroch vlákna, nazývaný koniec (-); A druhý koniec je opak, nazývaný koniec (+). Preto sa hovorí, že aktínové vlákno má polaritu.
Mnohokrát sú tieto komponenty známe ako mikrofilamenty, pretože sú zložkami cytoskeletu, ktoré vykazujú menší priemer.
Kde nájdeme Actin?
Actín je mimoriadne bežný proteín v eukaryotických organizmoch. Zo všetkých bunkových proteínov predstavuje aktín asi 5 až 10% - v závislosti od typu bunky. Napríklad v pečeni, každá z buniek, ktorá má takmer 5.108 Aktínové molekuly.
Charakteristika
Dve formy aktínu, monoméru a vlákna sa neustále nachádzajú v dynamickej rovnováhe medzi polymerizáciou a depolymérmi. Vo všeobecnosti existujú tri vlastnosti tohto javu:
1) Aktínové vlákna sú typické pre štruktúru svalového tkaniva a cytoskeletu eukaryotických buniek.
Môže vám slúžiť: 10 charakteristík najvýznamnejších lipidov2) Polymerizácia a depolimerizácia je regulovaný dynamický proces. Kde polymerizácia alebo agregácia monomérov aktínov G-ATP-MG+2 Stáva sa to na oboch koncoch. Že tento proces sa vyskytuje, závisí od podmienok média a regulačných proteínov.
3) Tvorba lúčov a relácií, ktoré tvoria aktínový cytoskelet, dáva silu motility buniek. Závisí to od proteínov, ktoré sa zúčastňujú na tvorbe krížových väzieb.
Funkcia
Svalová kontrakcia
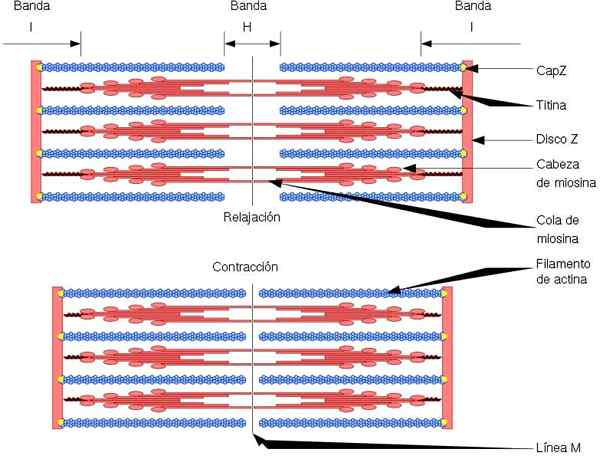
Funkčná a štrukturálna jednotka kostrového svalu je sarkomér, ktorý má dva typy vlákien: tenké vlákna, tvorené aktínmi a hrubými vláknami, tvorené myozínom. Obe vlákna sú organizované striedavo, presným geometrickým spôsobom. Povoliť kontrakciu svalov.
Tenké vlákna sú ukotvené do regiónov nazývaných disky z. Táto oblasť pozostáva z vláknovej siete, v ktorej sa nachádza proteín Capz, a na ktoré sú ukotvené konce (+) aktínových vlákien. Táto kotva zabraňuje koncovej depolerizácii (+).
Na druhej strane, na koncoch (-) aktínových vlákien sa nachádza tropomodulín a chráni ich pred depolymersiou. Okrem aktínu majú tenké vlákna tropomiozín a troponín, ktoré majú funkciu riadenia interakcií aktomiozínu.
Ako sa vyskytuje kontrakcia svalov?
Počas svalovej kontrakcie robia hrubé vlákna pohyby otáčania a vytiahnite tenké vlákna smerom do stredu sarkómu. To vytvára kĺzanie hrubých a tenkých vlákien.
Dĺžka hrubých a tenkých vlákien teda zostáva konštantná, ale zvyšuje prekrývanie medzi oboma vláknami. Dĺžka sarkoméru klesá ukotvením tenkých vlákien na disky Z.
Ako sa zastaví kontrakcia svalov?
ATP je energetická mena bunky. Preto je takmer vždy k dispozícii v živých svalových tkanivách. Berúc do úvahy vyššie uvedené mechanizmy, ktoré umožňujú relaxáciu svalov a zatknutie kontrakcií.
Dva proteíny, nazývané tropomiozín a troponín, majú v tomto fenoméne zásadnú úlohu. Tieto spolupracujú na blokovaní miest Myosínu odborov (čím sa bránia ich spojeniu s Actinom). Výsledkom je, že sval uvoľňuje.
Naopak, keď zviera zomrie. Osoba zodpovedná za toto tvrdenie tela je blokáda interakcie medzi myozínom a Actinom, krátko po smrti zvieraťa.
Jedným z dôsledkov tohto javu je potreba ATP na uvoľňovanie dvoch proteínových molekúl. Logicky nie je k dispozícii žiadna dostupnosť ATP v mŕtvych tkaninách a toto vydanie sa nemôže vyskytnúť.
Iné typy pohybu
Rovnaký mechanizmus, ktorý popisujeme (neskôr prehĺbime mechanizmus, ktorý je základom pohybu), sa neobmedzuje iba na svalové kontrakcie zvierat. Je zodpovedný za ameboidálne pohyby, ktoré pozorujeme v amébách a v niektorých koloniálnych formách.
Rovnakým spôsobom je cytoplazmatický pohyb, ktorý pozorujeme v rastlinách rias a pozemkov.
Regulácia polymerizácie a depolymerizácia aktínového vlákna
Kontrakcia tkaniva hladkého svalstva a buniek spôsobuje zvýšenie aktínu a zníženie aktív. Actínová polymerizácia sa vyskytuje v troch štádiách: 1) nukleacia, pomalý krok; 2) predĺženie, rýchly krok; a 3) stabilný stav. Rýchlosť polymerizácie sa rovná depolymerizácii rýchlosti.
Môže vám slúžiť: Fixistická teória: koncept, postuláty a autoriActínové vlákno rastie do konca (+) rýchlejšie ako koniec (-). Rýchlosť predĺženia je úmerná koncentrácii aktínových monomérov v rovnováhe s aktínovými vláknami, nazývanou kritická koncentrácia (CC).
CC pre koniec (+) je 0,1 um a pre koniec (-) je 0,8 um. To znamená, že na polymerizáciu konca (+) je potrebná 8 -krát menšia koncentrácia aktínových monomérov (+).
Actínová polymerizácia je regulovaná hlavne podvodmi s beta4 (TB4). Tento proteín spája aktínu a zachováva ho, čo bráni polymérne. Zatiaľ čo propylín stimuluje aktínovú polymerizáciu. Profil sa viaže na aktínové monoméry uľahčujúce polymerizáciu koncom (+), disociáciou komplexu aktínu-TB4.
Ďalšie faktory, ako je zvýšenie iónov (NA+, Klimatizovať+ alebo mg+2) uprednostňuje tvorbu vlákien.
Tvorba aktínového cytoskeletu
Tvorba aktínového cytoskeletu vyžaduje vypracovanie krížových väzieb medzi aktínovými vláknami. Tieto odkazy tvoria proteíny, ktorých vrcholy sú: majú domény Únie na Actin; Mnohé majú homológne domény s kalponínom; a každý typ proteínu je exprimovaný v určitom type bunky.
Vo Philopodes a stresových vláknach sú krížové putá medzi aktínovými vláknami vyrábané fascínom a Philamine. Tieto proteíny, respektíve, vytvárajú aktínové vlákna rovnobežné alebo majú rôzne uhly. Preto aktínové vlákna definujú tvar bunky.
Bunková oblasť s najvyšším množstvom aktínových vlákien sa nachádza v blízkosti plazmovej membrány. Táto oblasť sa nazýva kôra. Kortikálna cytoskelet je organizovaná v rôznych formách v závislosti od typu bunky a pripája sa k plazmatickej membráne fixáciou proteínov.
Niektoré z najlepšie opísaných cytoskeletónov sú svalové bunky, krvné doštičky, epitelové bunky a erytrocyty. Napríklad v svalových bunka. Tento komplex sa viaže na proteíny extracelulárnej matrice.
Model interakcie aktínu-miozín
Vedci vedci z lúčov navrhli štvorstupňový model na vysvetlenie interakcie aktínu a myozínu. Prvý krok sa vyskytuje s spojením ATP na myosínových hlavách. Táto únia vytvára konformačnú zmenu v proteíne, ktorá ho uvoľňuje z aktínu v malom vlákne.
Potom je ATP hydrolyzovaný na ADP, uvoľňuje anorganický fosfát. Molekula myozínu je ukotvená k novej podjednotke Actin, ktorá vytvára stav s vysokou energiou.
Uvoľňovanie anorganického fosfátu so sebou prináša zmenu myozínu, ktorá sa vracia k počiatočnej konformácii a pohyb malých vlákien, vzhľadom na hrubé vlákna, vzhľadom na hrubé vlákna, vzhľadom na silné vlákna. Toto hnutie spôsobuje pohyb oboch koncov sarkoméru a blíži sa k nim.
Posledný krok zahŕňa uvoľnenie ADP. V tomto okamihu je hlava myozínu zadarmo a môže sa pripojiť k novej molekule ATP.
Môže vám slúžiť: Biológia: História, aké štúdie, vetvy, základnePohyb buniek poháňaný aktínovou polymerizáciou
Motilita rektora je typ bunkovej pohyblivosti. Kroky tohto typu pohyblivosti sú: projekcia vedúcej osi adhézie k substrátu; adhézia na substrát; Zadné stiahnutie; a adhesie.
Projekcia vedúcej osi vyžaduje účasť proteínov, ktoré sa podieľajú na polymerizácii a depolimerizácii aktínových vlákien. Vedúca os sa nachádza v bunkovej kôre, ktorá sa nazýva lamellipodio. Kroky projekcie osi sú:
- Aktivácia receptora extracelulárnym signálom.
- Tvorba aktívnych GTPAS a fosfoinozitolu 4,5-bishoposfátu (PIP2).
- Aktivácia proteínu Wasp/Scar a Arp2/3, ktoré sa viažu na monoméry aktínu pri vytváraní dôsledkov vo vláknach aktínu.
- Rýchly rast aktínových vlákien, nakoniec zdobený myozínom, vetvenie. Membránový ťah sa vyrába vpred.
- Vyvrcholenie predlžovania produkovaného krycím proteínom.
- Hydrolýza ATP pripojená k aktínu v najstarších vláknach.
- Depolimerizácia aktínových vlákien podporovaných ADF/COFILINA.
- Výmena ADP pomocou ATP katalyzovaná profylaínou a generuje aktín G-ATP pripravený začať s predĺženými dôsledkami.
Choroby súvisiace s aktínom
Svalová dystrofia
Svalová dystrofia je degeneratívne ochorenie kostrového svalu. Je zdedený recesívne a je spojený s chromozómom X. Hlavne postihuje mužov s vysokou frekvenciou v populácii (jeden z 3.500 mužov). Matky týchto mužov sú asymptomatické heterozygotné a môžu chýbať rodinná anamnéza.
Existujú dve formy svalovej dystrofie, Duchenne a Becker a obe sú spôsobené defektmi génu dystrofínu. Tieto defekty pozostávajú z delécií, ktoré eliminujú axóny.
Dystrofín je proteín (427 kDa), ktorý tvorí krížové väzby medzi aktínovými vláknami. Má majstrovstvo únie aktínu v N-terminále a majstrovstvo membránovej únie v C-terminále. Medzi oboma doménami je tretia rúrková doména tvorená 24 tandemovými opakovaniami.
V svalovom kortikálnom retikule sa dystrofín podieľa na spojení aktínových vlákien do plazmatickej membrány prostredníctvom glykoproteického komplexu. Tento komplex sa tiež viaže na proteíny extracelulárnej matrice.
U pacientov, ktorí nemajú funkčný dystrofín, s Duchennovou svalovou dystrofiou, kortikálny cytoskelet nepodporuje plazmatickú membránu. V dôsledku toho je plazmatická membrána poškodená stresom opakovaných svalových kontrakcií.
Odkazy
- Devlin, T.M. 2000. Biochémia. Redakčné Reverté, Barcelona.
- Gunst, s.J., a Zhang, w. 2008. Actin cytoskeletálna dynamika v hladkom svale: Nová paradigma na reguláciu kontrakcie hladkého svalstva. Am J Physiol Cell Physiol, 295: C576-C587.
- Ubytovňa, h., Berk, a., Zipurski, s.L., Matsudaria, P., Baltimore, D., Darnell, J. 2003. Bunková a molekulárna biológia. Redakcia Medica Panamericana, Buenos Aires, Bogota, Caracas, Madrid, Mexiko, Sāo Paulo.
- Nelson, D. L., Cox, m. M. 2008. Lehninger-Principles of Biochemistry. W.H. Freeman, New York.
- Pfaendtner, j., Z kríža a. M., Voth, G. 2010. Prestavba aktínového vlákna pomocou aktínu depolymerizačný faktor/kofilín. Pnas, 107: 7299-7304.
- Pollard, T.D., Borisy, G.G. 2003. Bunková pohyblivosť poháňaná montážou a demontážou aktínových plnení. Cell, 112: 453-465.
- « Apolipoproteíny Čo sú, funkcie, typy
- Primárna štruktúra alkoholu, vlastnosti, nomenklatúra, príklady »