

Preklad procesu DNA v eukaryotách a prokaryotoch
- 2784
- 487
- Blažej Hrmo
Ten Preklad DNA Je to proces, ktorým informácie obsiahnuté v posloch vyrobených počas prepisu.
Z bunkového hľadiska je expresia génu relatívne komplexná hmota, ktorá sa vyskytuje v dvoch krokoch: transkripcia a translácia.
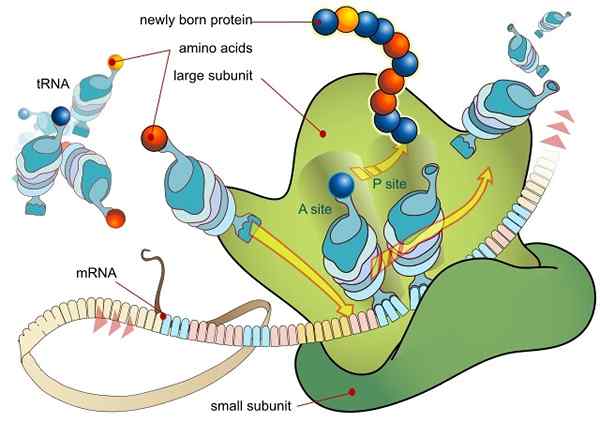
 Preklad RNA sprostredkovaný ribozómom (Zdroj: Ladyofhats / Public Domain, Via Wikimedia Commons)
Preklad RNA sprostredkovaný ribozómom (Zdroj: Ladyofhats / Public Domain, Via Wikimedia Commons) Všetky gény, ktoré sú exprimované (či už alebo nie sú kódujúce peptidové sekvencie, tj proteíny), spočiatku tak robia prenosom informácií obsiahnutých v jeho sekvencii DNA do Messenger MER Molecula (RNAM) procesom nazývaným transkripcia.
Prepis sa dosahuje špeciálnymi enzýmami známymi ako RNA polymeráza, ktoré používajú jeden z komplementárnych vlákien DNA génu ako formy pre syntézu molekuly „pred ANARNM“, ktorá je následne stíhaná, aby vytvorila zrelú zrelú zrelú zrelú zrelú.
Pre gény, ktoré kódujú proteíny, sú informácie obsiahnuté v zrelých RNM „čítané“ a preložené v aminokyselinách podľa genetického kódu, ktorý špecifikuje, ktorý kodón alebo nukleotidový trilet zodpovedá každej konkrétnej aminokyselinovej kyseline.
Špecifikácia aminokyselinovej sekvencie proteínu preto závisí od počiatočnej sekvencie dusíkových báz v DNA, ktorá zodpovedá génu a potom v RNA, ktorá transportuje uvedené informácie z jadra do cytosolu (v eukaryotických bunkách); Proces, ktorý je tiež definovaný ako syntéza proteínu riadenej RNM.
Vzhľadom na skutočnosť, že existuje 64 možných kombinácií 4 dusíkových báz, ktoré tvoria DNA a RNA a iba 20 aminokyselín, môže byť aminokyselina kódovaná rôznymi trojicami (kodóny), takže sa hovorí, že genetický kód je “ degenerovať “(s výnimkou aminokyselinového metrií, ktorý je kódovaný jedinečným AUG kodónom).
[TOC]
Preklad Eukaryota (kroky procesov)
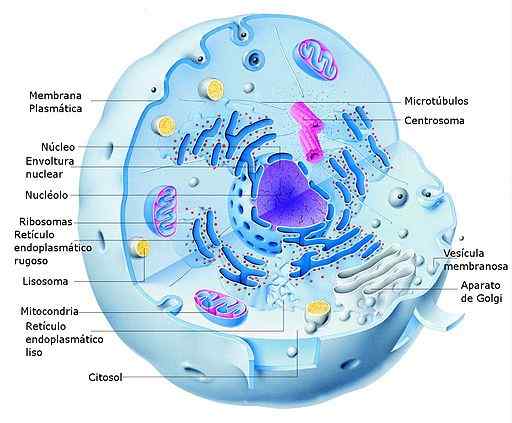
Schéma bunky Eukaryot Animal Eukaryot a jej častí (zdroj: Alejandro Porto [CC0] cez Wikimedia Commons) V eukaryotických bunkách sa transkripcia uskutočňuje v jadre a translácii do cytosolu, takže RNM, ktoré sa tvoria počas prvého procesu ).
Je dôležité spomenúť, že rozdelenie transkripcie a translácie v eukaryotoch platí pre jadro, ale nie je to isté pre organely s ich vlastným genómom, ako sú chloroplasty a mitochondrie, ktoré majú viac podobných systémov ako prokaryotické organizmy.
Eukaryotické bunky majú tiež cytosolické ribozómy viazané na membrány endoplazmatického retikula (hrubý endoplazmatický retikula), v ktorom sa translácia proteínov, ktoré majú byť vložené do bunkových membrán alebo ktoré vyžadujú post -translačné spracovanie, ktoré sa vyskytuje v uvedenom kompartmente.
- RNM spracovanie pred ich prekladom
RNM sú upravené na svojich koncoch, keď sú prepisované:
- Keď sa 5 'koniec RNM objaví z povrchu RNA polymerázy II počas transkripcie, je to „okamžite napadnuté“ skupinou enzýmov, ktoré syntetizujú „kapucň“ zloženú zo 7-metyl guanilado a ktoré je spojené s terminálom Nukleotid RNM prostredníctvom 5 'triffosfátového spojenia, 5'.
Môže vám slúžiť: kodón- 3 'koniec RNM trpí „klivaje“ endonukleázou, ktorá generuje bezplatnú hydroxylovú skupinu 3, ktorá sa spája s „ristrou“ alebo „chvostom“ adenínového odpadu (od 100 do 250), ktoré sa pridávajú rovnaké v rovnakom čas na enzým Poly A) polymeráza.
„Kapucňa 5“ a „chvost Poly A “spĺňajú funkcie pri ochrane molekúl RNM proti degradácii a navyše pracujú na transporte zrelých transkriptov do cytosolu a pri začatí a ukončení prekladu.
COrte a empalme
Po transkripcii prechádzajú „primárne“ RNM s dvoma modifikovanými extrémami, ktoré sú stále prítomné v jadre, prechádzajú procesom „rezania a zostrihu“, pomocou ktorého sú intronické sekvencie všeobecne eliminované a výsledné exóny sú spojené (postregionálne spracovanie), s tým, čo sa získajú zrelé prepisy, ktoré opustia jadro a dosahujú cytosol.
Rez a zostrih je vykonávané riboproteickým komplexom nazývaným Espliceleosóm (Anglicizmus Zostrih), tvorené piatimi malými ribonukleoproteínmi a molekulami RNA, ktoré sú schopné „rozpoznať“ oblasti, ktoré musia byť eliminované z primárneho prepisu.
V mnohých eukaryotoch je fenomén známy ako „rezanie a alternatívny kĺb“, čo znamená, že rôzne typy post -registračných modifikácií môžu spôsobiť rôzne alebo izoenzýmové proteíny, ktoré sa navzájom líšia v niektorých aspektoch svojich sekvencií.
- Ribozómy
Keď zrelé transkripty opúšťajú jadro a sú transportované na transláciu do cytosolu, sú spracované translačným komplexom známym ako ribozóm, ktorý pozostáva z proteínového komplexu spojeného s molekulami RNA.
Ribozómy sa skladajú z dvoch podjednotiek, jednej „veľkej“ a druhej „malej“, ktoré sú voľne disociované v cytosóle a spájajú sa alebo sa pridávajú k molekule mRNA, ktorá sa prekladá.
Spoja medzi ribozómami a mRNA závisí od špecializovaných molekúl RNA, ktoré sú spojené s ribozomálnymi proteínmi (ribozomálna RNA alebo RNA a prenos alebo Arnt RNA), z ktorých každý vykonáva špecifické funkcie.
ARNT sú molekulárne „adaptéry“, pretože prostredníctvom jedného z ich koncov môžu „čítať“ každý kodón alebo triplet v zrelej RNA (komplementaritou báz) a cez druhý sa môžu pripojiť k aminokyselinám kódovanej kodónom „čítaním“.
Na druhej strane molekuly RNR sú zodpovedné za urýchlenie (katalyzovanie) väzbový proces každej aminokyseliny v rodnom peptidovom reťazci.
Eukaryotický zrelý RNM môže byť „prečítaný“ mnohými ribozómami, čo mnohokrát, ako to naznačuje bunka. Inými slovami, rovnaké RNM môže viesť k mnohým kópiám toho istého proteínu.
Počiatočný kodón a čítací rámec
Keď sa k zrelému RNM priblíži k ribozomálnym podjednotkám, riboproteický komplex „sken“ Sekvencia uvedenej molekuly, až kým nezistí štartovací kodón, čo je vždy AUG a naznačuje zavedenie zvyšku metionínu.
Môže vám slúžiť: Monoploidy: Ako sa to deje, organizmy, frekvencia a užitočnosťKodón AUG definuje čítací rámec pre každý gén a navyše definuje prvú aminokyselinu všetkých proteínov preložených do prírody (táto aminokyselina je často eliminovaná posttranslačným).
Kodóny
Boli identifikované tri ďalšie kodóny, ktoré vyvolávajú ukončenie prekladu: UAA, UAG a UGA.
Mutácie, ktoré naznačujú zmenu dusíkových báz v triplete, ktorý kóduje aminokyselinu a vedú k koncovému kodónom, sú známe ako nezmyselné mutácie, pretože spôsobujú predčasné zadržiavanie procesu syntézy, ktorý tvorí kratšie kratšie proteíny.
Nepreložené regióny
V blízkosti 5 'konca zrelých molekúl RNM existujú regióny, ktoré nie sú preložené (UTR z angličtiny Netranslate región), tiež nazývané sekvencie „Leader“, ktoré sa nachádzajú medzi prvým nukleotidom a začiatkom prekladu (AUG).
Tieto oblasti UTR, ktoré nie sú preložené, majú špecifické miesta pre spojenie s ribozómami a ľuďmi, napríklad majú približnú dĺžku 170 nukleotidov, medzi ktorými existujú regulačné oblasti, miesta väzby na proteíny, ktoré pracujú pri regulácii translácie atď.
- Začiatok prekladu
Preklad, ako aj transkripcia pozostávajú z 3 fáz: jedna zo zasvätenia, ďalšia predĺženie a nakoniec jedno z ukončenia.
Zasvätenie
Skladá sa zo zhromaždenia translačného komplexu na RNM, ktorý si zaslúži spojenie troch proteínov známych ako iniciačné faktory (ak angličtinu Iniciačný faktor) If1, if2 a if3 k malej podjednotke ribozómu.
Komplex „pred -iniciácie“ tvorený iniciačnými faktormi a malá ribozomálna podjednotka je zase spojená s ARNT, ktorý „načíta“ zvyšok meonínu a táto sada molekúl sa viaže na RNAM, blízko štartovacieho kodónu AUG.
Tieto udalosti vedú k zväzku RNM s veľkou ribozomálnou podjednotkou, ktorá vedie k uvoľňovaniu iniciačných faktorov. Veľká podjednotka ribozómu má 3 miesta únie pre molekuly ARNT: miesto A (aminokyselina), miesto P (polypeptid) a miesto E (výstup).
Stránka A sa spája s aminoacil-Arnt Anticod, ktorý sa dopĺňa k prekladaniu mRNA; Miesto p je miesto, kde sa aminokyselina prenáša z ARNT do rodiaceho sa peptidu a miesto S je miesto, kde je v Arnt „prázdne“ a potom sa uvoľní do cytosolu po dodaní aminokyseliny.
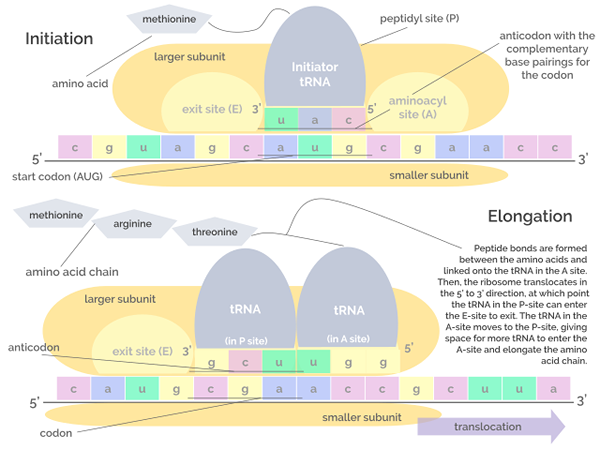 Grafické znázornenie fáz iniciácie a predĺženia prekladu (Zdroj: Jordan Nguyen/CC By-S (https: // creativeCommons.Org/licencie/By-SA/4.0) Via Wikimedia Commons)
Grafické znázornenie fáz iniciácie a predĺženia prekladu (Zdroj: Jordan Nguyen/CC By-S (https: // creativeCommons.Org/licencie/By-SA/4.0) Via Wikimedia Commons) Predĺženie
Táto fáza pozostáva z „pohybu“ ribozómu pozdĺž molekuly mRNA a preklad každého kodónu, ktorý „číta“, čo znamená rast alebo predĺženie polypeptidového reťazca pri narodení.
Tento proces si vyžaduje faktor známy ako predlžovací faktor GTP vo forme GTP, ktorý pri prekladaní molekuly RNM poháňa translokáciu predlžovacích faktorov pozdĺž molekuly RNM.
Môže vám slúžiť: fragmenty okazakiAktivita prenosázy peptidil ribozomálnych RNA umožňuje tvorbu peptidových väzieb medzi následnými aminokyselinami, ktoré sa pridávajú do reťazca.
Ukončenie
Preklad končí, keď ribozóm spĺňa niektorý z koncových kodónov, pretože ARN tieto kodóny nerozpoznávajú (nekódujú aminokyseliny). Sú spojené aj proteíny známe ako faktory uvoľňovania, ktoré uľahčujú oddelenie ribosoma MR.
PROCARY PREKLAD (Passes-proceses)
V prokaryotoch, rovnako ako v eukaryotických bunkách, sa ribozómy zodpovedné syntézou proteínov nachádzajú v cytosóle (čo je tiež pravda pre transkripčné mechanizmy), skutočnosť, ktorá umožňuje rýchle zvýšenie cytosolovej koncentrácie proteínu pri expresii génov, ako kóduje, zvyšuje sa.
Aj keď v týchto organizmoch to nie je mimoriadne bežný proces, primárny RNM produkovaný počas transkripcie môže trpieť post -registračným dozrievaním prostredníctvom „strihu a zostrihu“. Najbežnejším je však pozorovať ribozómy pripojené k primárnym transkribovaným, ktoré ho prekladajú v rovnakom čase, ako sa prepisuje zo zodpovedajúcej sekvencie DNA.
Vzhľadom na vyššie uvedené sa preklad v mnohých prokaryotoch začína koncom 5 ', pretože 3' koniec mRNA zostáva spojený s DNA formou (a vyskytuje sa súbežne s transkripciou).
Nepreložené regióny
Prokaryotické bunky tiež produkujú RNM s netranslačnými oblasťami, ktoré sú známe ako „Shine-Dalgarno Box“ a ktorých konsenzuálna sekvencia je AgGagg. Ako je zrejmé, oblasti baktérií UTR sú podstatne kratšie ako oblasti eukaryotických buniek, hoci počas translácie vykonávajú podobné funkcie.
Spracovanie
V baktériách a iných prokaryotických organizmoch je translačný proces dosť podobný procesu eukaryotických buniek. Skladá sa tiež z troch fáz: iniciácia, predĺženie a ukončenie, ktoré závisia od špecifických prokaryotických faktorov, ktoré sa líšia od tých, ktoré používajú eukaryoty.
Napríklad predĺženie závisí od predlžovacích faktorov známych ako EF-TU a EF-TS namiesto eukaryotického faktora G.
Odkazy
- Alberts, b., Johnson, a., Lewis, J., Raff, m., Roberts, K., & Walter, P. (2007). Biológia molekulárnej bunky. Girlandská veda. New York, 1392.
- Clay, s. & Hnedá, w. (2008) Translácia: DNA na mRNA na proteín. Prírodné vzdelávanie 1 (1): 101.
- Griffiths, a. J., Wessler, s. R., Lewontin, r. C., Gelbart, w. M., Suzuki, D. Tón., & Miller, J. H. (2005). Úvod do genetickej analýzy. Macmillan.
- Ubytovňa, h., Berk, a., Kaiser, C. Do., Krieger, m., Scott, m. P., Bretscher, a.,… & Matsudaira, str. (2008). Biológia molekulárnych buniek. Macmillan.
- Nelson, D. L., Lehninger, a. L., & Cox, M. M. (2008). Lehninger princípy biochémie. Macmillan.
- Rosenberg, L. A., & Rosenberg, D. D. (2012). Ľudské gény a genómy: veda. Zdravie, spoločnosť, 317-338.
- « Charakteristiky Sorbus ária, biotop, vlastnosti, kultúra
- Boletus aereus funkcie, biotop, identifikácia, recepty »