

Syntéza typov lipidov a ich hlavné mechanizmy
- 4064
- 1306
- Denis Škriniar
Ten Syntéza lipidov Skladá sa zo série enzymatických reakcií, pomocou ktorých sa uhľovodíky s krátkym reťazcom kondenzujú za vzniku molekúl s dlhším reťazcom, ktoré môžu následne trpieť rôznymi chemickými modifikáciami.
Lipidy sú veľmi rozmanitý druh biomolekúl syntetizovaných všetkými živými bunkami a špecializujú sa na viaceré základné funkcie na udržiavanie životnosti buniek.
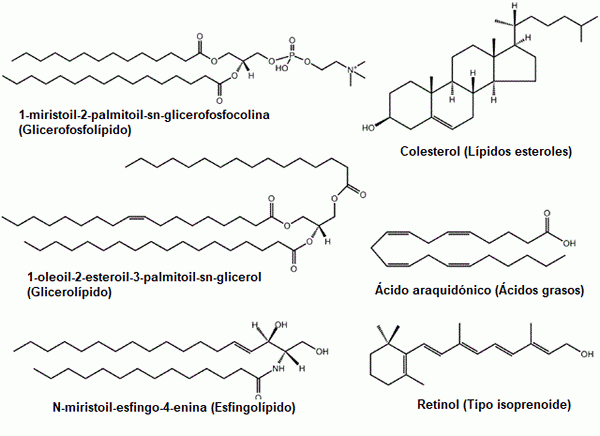 Niektoré príklady bežných lipidov: glycerofosfolipidy, steroly, glycerolipidy, mastné kyseliny, sfingolipidy a prenooly (zdroj: Pôvodný Uploader bol LMAP v anglickej Wikipedia. / Gfdl 1.2 (http: // www.GNU.Org/licencie/staré licenky/FDL-1.2.html) cez Commons, upravené Raquel Parada)
Niektoré príklady bežných lipidov: glycerofosfolipidy, steroly, glycerolipidy, mastné kyseliny, sfingolipidy a prenooly (zdroj: Pôvodný Uploader bol LMAP v anglickej Wikipedia. / Gfdl 1.2 (http: // www.GNU.Org/licencie/staré licenky/FDL-1.2.html) cez Commons, upravené Raquel Parada) Lipidy sú hlavnými zložkami biologických membrán, čo z nich robí základné molekuly pre existenciu buniek ako izolovaných entít ich okolia.
Niektoré lipidy majú tiež špecializované funkcie, ako sú pigmenty, kofaktory, transportéry, detergenty, hormóny, intra-extracelulárne poslovia, kovalentné kotvy pre membránové proteíny atď. Preto je schopnosť syntetizovať rôzne typy lipidov pre prežitie všetkých živých organizmov prežitie všetkých živých organizmov.
Táto veľká skupina zlúčenín je tradične klasifikovaná do niekoľkých kategórií alebo podskupín: mastné kyseliny (nasýtené a nenasýtené), lipidy glyceridov (lipoproteíny) (lipoproteíny).
[TOC]
Typy lipidov a ich hlavné mechanizmy syntézy
Všetky reakčné sekvencie trasy biosyntézy lipidov sú endogonické a redukčné. Inými slovami, všetky používajú ATP ako zdroj energie a znížený elektrónový transportér, ako je NADPH, ako redukčný výkon.
Ďalej budú opísané hlavné reakcie biosyntetických trás hlavných typov lipidov, tj mastných a eikozanoidných kyselín, triacylglycerolov a fosfolipidov a sterolov (cholesterol) (cholesterol) (cholesterol).
- Syntéza mastných kyselín
Mastné kyseliny sú mimoriadne dôležité molekuly z hľadiska lipidov, pretože sú súčasťou najrelevantnejších lipidov v bunkách. Jeho syntéza, na rozdiel od toho, čo si mnohí vedci mysleli počas prvých štúdií, nepotýka opačnej cesty ich β-oxidácie.
V skutočnosti sa táto metabolická cesta vyskytuje v rôznych bunkových kompartmentoch a vyžaduje účasť sprostredkovateľa troch atómov uhlíka známych ako Malonyl-CoA, čo nie je potrebné pri oxidácii.
 Maonyl-coa. Neuroteker / krčma doména
Maonyl-coa. Neuroteker / krčma doména Okrem toho úzko súvisí so sulfhydrilnými skupinami proteínov známych ako transportéry skupiny Acyl (ACP, angličtina Proteíny s acylovým nosičom).
Všeobecne platí, že syntéza mastných kyselín, najmä v prípade dlhého reťazca, je postupný proces, v ktorom sa v každom „návrate“ opakujú štyri kroky, a počas každého návratu je skupina nasýtenej kyseliny, ktorá je substrátom pre ďalší, ktorý naznačuje ďalšiu kondenzáciu s novou molekulou Malonyl-CoA.
Pri každom zákrute alebo reakčnom cykle reťazec mastných kyselín rozširuje dva uhlíky, až kým nedosiahne dĺžka 16 atómov (palmitát), po ktorom cyklus opúšťa.
Tvorba Malonyl-CoA
Tento sprostredkovateľ troch atómov uhlíka je ireverzibilne tvorený z acetyl-CoA vďaka pôsobeniu acetyl-CoA karboxylázového enzýmu, ktorý má skupinu prostaty biotínu, ktorá je kovalentne spojená s enzýmom a ktorá sa podieľa na tejto katalýze v tejto katalýze v dvoch krokoch.
V tejto reakcii sa karboxylová skupina odvodená z bikarbonátovej molekuly (HCO3-) prenáša do biotínu vo forme závislej od ATP, kde biotinylová skupina plní funkciu „dočasného dopravníka“ molekuly, zatiaľ čo prenosy do acetyl-CoA, výroba Malonyl-CoA.
Môže vám slúžiť: oxidázový test: základ, postup a použitieV sekvencii syntézy mastných kyselín použité redukčné činidlo je NADPH a aktivačné skupiny sú dve skupiny TIOL (-SH), ktoré sú súčasťou multienzimatického komplexu nazývaného mastné kyseliny syntázy, ktorá je najdôležitejšia pri syntetickej katalýze syntetickej katalýzy.
U stavovcov je komplex mastných kyselín syntázy súčasťou jediného veľkého polypeptidového reťazca, v ktorom je zastúpených 7 enzymatických aktivít charakteristických pre cestu syntézy, ako aj hydrolytická aktivita potrebná na uvoľňovanie sprostredkovateľov na konci syntézy.
 Štruktúra enzýmov mastných kyselín Syntasa (zdroj: Boehringer Ingelheim/CC By-SA (https: // creativeCommons.Org/licencie/By-SA/4.0) Via Wikimedia Commons)
Štruktúra enzýmov mastných kyselín Syntasa (zdroj: Boehringer Ingelheim/CC By-SA (https: // creativeCommons.Org/licencie/By-SA/4.0) Via Wikimedia Commons) Sedem enzymatických aktivít tohto komplexu sú: transportný proteín skupiny ACILO (ACP), acetyl-CoA-ACP transacetilas cetoacil-ACP reduktáza (KR), β-hydroxyacil-ACP dehydratasa (HD) a REGGER-ACP reduktáza (ER) (ER).
Predtým, ako sa môžu vyskytnúť kondenzačné reakcie na zostavenie reťazca mastných kyselín, sú dve skupiny tiolov v enzymatickom komplexe „naložené“ do acylových skupín: v prvom rade sa acetyl -coA prenesie do skupiny cysteine -sh. V p-cethoacyl-ACP syntáza komplexu, reakcia katalyzovaná acetyl-CoA-ACP transacetyla enzýmom (AT) (AT).
Následne sa malonická skupina prenáša z molekuly Malonyl -CoA do skupiny -SH skupiny ACILO Group (ACP) dopravníka ACILO (ACP) je súčasťou syntázy komplexu mastných kyselín.
Sekvencia štyroch reakcií pre každý „návrat“ reakčného cyklu je nasledovná:
- Kondenzácia: skupiny acetyl a malonyl „naložené“ v enzýme sa kondenzujú a vytvárajú molekulu acetoacetyl -ACP, ktorá je spojená s časťou AKT prostredníctvom skupiny -SH. V tomto kroku je molekula CO2 a je katalyzovaná syntázou β-zoacyl-ACP (acetylová skupina zaberá polohu „terminálneho metylu“ komplexu acetoacetyl-ACP).
- Redukcia karbonylovej skupiny: Karbonylová skupina v polohe C3 acetoacetyl-ACP sa redukuje na vytvorenie D-P-hydroxibutiríl-ACP, reakcia katalyzovaná β-cethoacyl-ACP reduktáza, ktorú NADPH používa ako elektrónový daror.
- Dehydratácia: C2 a C3 uhlíky D-P-hydroxibutiríl-ACP nemajú molekuly vody, čo tvoria dvojitú väzbu, ktorá končí výrobou novej zlúčeniny previesť-∆2-butenoil-ACP. Tento proces je sprostredkovaný enzýmom pydroxyacyl-ACP dehydratázy (HD) (HD).
- Dvojitá redukcia väzby: Dvojitá väzba zlúčeniny vytvorenej v kroku dehydratácie je nasýtená (redukovaná), aby sa vznikla na butiril-ACP pomocou reakcie katalyzovaného enzýmovým hnevom reduktázou (ER), ktorá tiež používa NADPH ako redukčné činidlo ako redukčné činidlo.
Syntéza reakcií sa vyskytuje, až kým sa nevytvorí molekula palmitátu (16 atómov uhlíka), ktorá sa hydrolyzuje z enzymatického komplexu a uvoľňuje sa ako možný prekurzor mastných kyselín reťazcov s väčšou dĺžkou, ktoré sa produkujú elongačnými systémami mastných kyselín umiestnených v hladkom hladkom časť endoplazmatického retikula a v mitochondriách.
Môže vám slúžiť: fauna a flóra peruánskeho moraOstatné modifikácie, ktoré tieto molekuly môžu trpieť, napríklad deaturácie, sa katalyzujú rôznymi enzýmami, ktoré sa zvyčajne vyskytujú v hladkom endoplazmatickom retikule.
- Syntéza eikosanoidov
Eikosanoidy sú bunkové lipidy, ktoré majú funkcie, ako sú molekuly posolov „krátky“, produkované niektorými tkaninami na komunikáciu so svojimi susednými látkami. Tieto molekuly sú syntetizované z polynenasýtených mastných kyselín 20 atómov uhlíka.
Prostaglandíny
V reakcii na hormonálny stimul fosfolipázový enzým útočí na membránové fosfolipidy a uvoľňuje araquidonato z glycerolu uhlíka 2. Táto zlúčenina sa premieňa na prostaglandíny vďaka enzýmu hladkého endoplazmatického retikula s bifunkčnou aktivitou: cyklooxygenáza (COX) alebo prostaglandín H2 syntázu.
Tromboxan
Prostaglandíny sa dajú premieňať na tromboxans vďaka tromboxanu synlása v krvných krvných doštičkách (trombocyty). Tieto molekuly sa podieľajú na počiatočných krokoch koagulácie krvi.
- Syntéza triacylglycerolov
Mastné kyseliny sú základné molekuly pre syntézu iných komplexnejších zlúčenín v bunkách, ako sú triacylglyceroly alebo lipidová glycerofosfolipidová lipidová membrána (procesy, ktoré závisia od bunkových metabolických potrieb).
Zvieratá produkujú triacylglycelály a glyceofosfalipidy z dvoch bežných prekurzorov: acyl-coA a 3-fosfát L-glycerol. Acyl-coA acyl sa vyrába acyl-CoA syntézami, ktoré sa podieľajú na β-oxidácii, zatiaľ čo 3-fosfát L-glycerol sa získa z glykolýzy a účinkom dvoch alternatívnych enzýmov: 3-fosfát glycerol dehydrogenáza a kináza glycercercercercercercercercercercercercercercerol a kinázový glycercercercercel.
Triacylglyceroly sú tvorené reakciou medzi dvoma molekulami flo-CoA aCIL a 3-fosfátovou diacylglycerolovou molekulou; Tieto prenosové reakcie sú katalyzované špecifickými prenosmi.
V tejto reakcii je spočiatku kyselina fosfatidová, ktorá je defosforylovaná fosfatázou kyseliny enzýmovej fosfatidovej kyseliny za vzniku 1,2 diacylglycerolu, ktorý je opäť schopný prijať tretiu molekulu tukovo-coA acyl, produkujúcej triacilglycerolol.
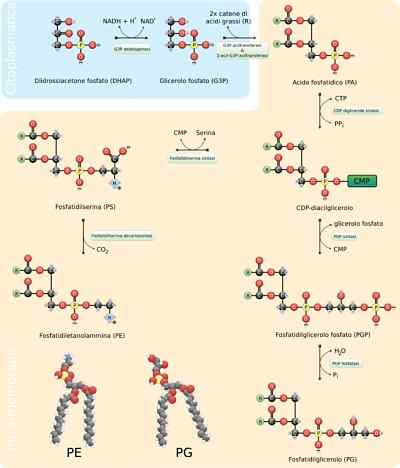
- Syntéza fosfolipidov
Fosfolipidy sú extrémne variabilné molekuly, pretože mnohé rôzne sa môžu tvoriť kombináciou mastných kyselín a skupín „hlavy“ s glycerolovým kostrom (glyceofosfolipidy) alebo sfhinozínom (sfingolipidy), ktoré ich charakterizujú.
Valné zostavenie týchto molekúl vyžaduje syntézu kostry glycerolu alebo sférického spojenia so zodpovedajúcimi mastnými kyselinami, buď sterifikáciou alebo amidáciou, pridanie hydrofilnej skupiny „hlavy“ prostredníctvom fosfodiérovej väzby a , zmena alebo výmena týchto posledných skupín.
V eukaryotoch sa tento proces vyskytuje v hladkom endoplazmatickom retikule a tiež v vnútornej mitochondriálnej membráne, kde môžu zostať neurčito alebo odkiaľ sa môžu translokovať na iné miesta.
Reakčné kroky
Prvé kroky syntézy reakcie glycerofosfolipidov sú ekvivalentné s produkciou triacylglycerolov, pretože molekula glycerolu 3-fosfátu je esterifikovaná na dve molekuly mastných kyselín v uhlíkoch 1 a 2, tvoriaci. Je bežné nájsť fosfolipidy, ktoré majú nasýtené mastné kyseliny v C1 a nenasýtené v C2 glycerolu.
Môže vám slúžiť: rezistinaKyselina fosfatidová sa môže tiež produkovať fosforyláciou diacylglycerolovej molekuly už syntetizovanej alebo „recyklovanej“.
Polárne skupiny týchto molekúl „hlavy“ sa tvoria prostredníctvom fosfodiérových väzieb. Prvá vec, ktorá by sa mala stať, aby bol tento proces správne.
Ak sa táto molekula viaže na diacylglycerol, vytvorí sa CDP-Diacylglycerol („aktivovaný“ tvar kyseliny fosfatidovej), ale môže sa to stať aj v hydroxylovej skupine skupiny „hlava“.
Napríklad v prípade fosfatidylserínu sa diacglycerol aktivuje kondenzáciou molekuly kyseliny fosfatidovej s molekulami cytikuly trif-citidínu (CTP), tvoriacou CDP-diacylglycerol a eliminuje pyrofosfát pyrofosfátu.
Ak sa molekula MMP (cytidín monofosfát) pohybuje nukleofilným aktónom serínu alebo hydroxyl hydroxylu v uhlíku 1 3-fosfátového glycerolu. Fosfáty produkujú fosfatidylglycerol.
Obe molekuly produkované týmto spôsobom slúžia ako prekurzory iných membránových lipidov, ktoré často zdieľajú biosyntetické trasy od seba navzájom.
- Syntéza cholesterolu
Cholesterol je nevyhnutnou molekulou pre zvieratá, ktoré môžu byť syntetizované jeho bunkami, takže v dennej diéte nie je nevyhnutná. Táto molekula 27 atómov uhlíka sa vyrába z predchodcu: acetát.
Táto komplexná molekula je tvorená z acetyl-CoA v štyroch hlavných fázach:
- Kondenzácia troch acetátových jednotiek za vzniku mevalonata, sprostredkovateľskej molekuly 6 uhlíkov (najprv sa vytvára molekula acetoacetyl-CoA s dvoma acetyl-CoA (HMG-enzým) a potom ďalším z phydroxy-p-metylglutaril-coA (HMG-enzým) COA) (HMG-COA synteta enzým). Mevalonato je vytvorený z HMG-COA a vďaka enzýmu HMG-COA reduktázy.
- Mevalonato konverzia na izoprénové jednotky. Prvé 3 fosfátové skupiny sa prenášajú z 3 molekúl ATP na mevalonato. Jeden z fosfátov sa stratí spolu so susednou karbonylovou skupinou a je vytvorený
- Polymerizácia alebo kondenzácia 6 izoprénových jednotiek 5 atómov uhlíka za vzniku 30 -karbonových atómov (lineárna molekula) HOLE (lineárna molekula).
- Cicration of the Escualana za vzniku 4 kruhov steroidného jadra cholesterolu a následné chemické zmeny: oxidácie, migrácia a eliminácia metylových skupín atď., Čo poskytuje cholesterol.
Odkazy
- Garrett, R. H., & Grisham, C. M. (2001). Zásady biochémie: s ľudským zameraním. Vydavateľstvo Brooks/Cole.
- Murray, r. Klimatizovať., Granner, D. Klimatizovať., Mayes, P. Do., & Rodwell, V. W. (2014). Harperova ilustrovaná biochémia. McGraw-Hill.
- Nelson, D. L., Lehninger, a. L., & Cox, M. M. (2008). Lehninger princípy biochémie. Macmillan.
- Jacquemyn, J., Cascaho, a., & Goodchild, r. A. (2017). Ins a výstupy endoplazmatickej biosyntézy lipidov kontrolovanej retikula. Embo Reports, 18 (11), 1905-1921.
- Ohlrogge, j., & Prezrite, J. (Devätnásť deväťdesiatpäť). Biosyntéza lipidov. Rastlinná bunka, 7 (7), 957.
- « Životopis, vláda a príspevky José López Portillo
- Charakteristiky tropickej džungle v Mexiku, umiestnenie, reliéf, flóra, fauna »