

SGLT (Transportné proteíny transportu sodného a glukózy)
- 4217
- 383
- Tomáš Mydlo
Ten Transportné proteíny sodíka a glukózy (Sglt) Sú zodpovední za vykonanie aktívneho transportu glukózy v cicavčích bunkách proti koncentračnému gradientu. Energia potrebná na to, aby bola táto preprava možná, je získaná z kotransportu sodného rovnakým smerom (Simport).
Jeho umiestnenie je obmedzené na membránu buniek, ktoré tvoria epitelové tkanivá zodpovedné za absorpciu a reabsorpciu živín (tenké črevo a proximálny tvarovaný tubulo obličiek).
K dnešnému dňu bolo opísaných iba šesť izoforiem patriacich do tejto rodiny transportérov: SGLT-1, SGLT-2, SGLT-3, SGLT-4, SGLT-5 a SGLT-6. Vo všetkých z nich elektrochemický prúd generovaný transportom sodného iónu poskytuje energiu a indukuje konformačnú zmenu v štruktúre potrebného proteínu na translokovanie metabolitu na druhej strane membrány.
Všetky tieto izoformy sa však navzájom líšia prezentáciou rozdielov v:
- Stupeň afinity, ktorú majú pre glukózu,
- Schopnosť vykonávať transport glukózy, galaktózy a aminokyselín,
- Stupeň, do akej sú inhibované Florizinou a
- Umiestnenie tkaniva.
[TOC]
Molekulárne mechanizmy transportu glukózy
Glukóza je monosacharid šiestich atómov uhlíka, ktorý používa väčšina existujúcich typov buniek na získanie energie prostredníctvom metabolických oxidačných dráh.
Vzhľadom na svoju veľkú veľkosť a v podstate hydrofilitnú povahu nie je schopný prekročiť bunkové membrány voľnou difúziou. Jeho mobilizácia cytosolu preto závisí od prítomnosti transportných proteínov v týchto membránach.
Doteraz študované transportéry glukózy vykonávajú transport tohto metabolitu pasívnymi alebo aktívnymi transportnými mechanizmami. Pasívna doprava sa líši od aktívneho transportu, v ktorom nevyžaduje vykonávanie dodávok energie, pretože sa vyskytuje v prospech koncentračného gradientu.
Môže vám slúžiť: epidermis cibuleProteíny zapojené do pasívneho transportu glukózy patria do rodiny šírených diseminovaných transportérov, ktoré sa nazývajú skratkou v angličtine pojmu „transportéry glukózy“. Zatiaľ čo tí, ktorí vykonávajú aktívny transport, sa nazývali SGLT „transportnými proteínmi sodíka-glukóza“.
Posledne menovaná získava potrebnú voľnú energiu na vykonanie transportu glukózy proti jej koncentračnému gradientu kotransportu iónov sodného. Bolo identifikovaných najmenej 6 izoforiem SGR a zdá sa, že jeho poloha je obmedzená na membrány epitelových buniek.
Charakteristika SGLT
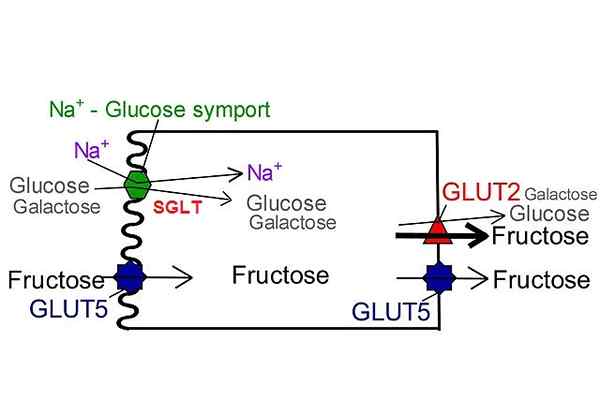
Ako je uvedené podľa názvu, transportné proteíny sodíka-glukóza vykonávajú typ sodíka a glukózy alebo sodíka a sodíka a galaktózy cytosolu do bunkového cytosolu.
Transport sodíka v prospech koncentračného gradientu je zodpovedný za vytvorenie nevyhnutnej voľnej energie na mobilizáciu glukózy z zóny s nízkou koncentráciou do vysokej koncentrácie.
Tieto transportéry sú súčasťou skupiny anti -transportu a sympatie sodíka a vodíka, ktorých gény nie sú fylogeneticky príbuzné.
Jeho umiestnenie je obmedzené na luminálnu membránu epitelových buniek tkanív zapojených do absorpcie živín, ako je tenké črevo a proximálny tvarovaný tubulu.
Môže vám slúžiť: Coxiella BurnetiiTypy SGLT
K dnešnému dňu bolo identifikovaných šesť transportérov SGLT (SGLT-1, SGLT-2, SGLT-3, SGLT-4, SGLT-5 a SGLT-6). Všetky ukazujú rozdiely v preferencii transportu glukózy alebo galaktózy, afinity, ktorú prezentujú pre tieto cukry a sodík, a skutočnosť, že Florizina môže byť inhibovaná.
SGLT1 má schopnosť transportovať galaktózu okrem glukózy s veľmi podobnou kinetikou, zatiaľ čo SGLT2 iba transportuje glukózu.
GSLT štruktúra
Dĺžka peptidovej sekvencie SGLT proteínov sa pohybuje od 596 do 681 aminokyselinových odpadov. Medzitým sa homologické percentá medzi sekvenciami pohybujú medzi 50 a 84 % vo vzťahu k SGLT-1, čo je najväčší stupeň divergencie v sekvenciách, ktoré zodpovedajú extracelulárnej amino-termináli a terminálu karboxyl.
Všeobecná štruktúra sa zhoduje s štruktúrou multišo integrálneho proteínu, to znamená, že prechádza cez membránu viackrát cez transmembránové domény bohaté na hydrofóbne aminokyseliny.
Tieto transmembránové domény sú celkom 14 a každá z nich má sekundárnu štruktúru a-helix. Segmenty 1, 2, 5, 7 a 9 sú priestorovo usporiadané v centrálnej polohe, ktorá vedie k tvorbe hydrofilných pórov.
Prostredníctvom hydrofilných tranzitov pórov glukózy, ako aj ktorýkoľvek z ďalších metabolitov, pre ktoré môže transportér mať afinitu. Zostávajúce špirálové fragmenty sú usporiadané paralelne, aby sa kanálovi poskytlo väčšiu štrukturálnu stabilitu.
Funkcie SGLT
Členovia rodiny SGLT nevykazujú pre glukózu jedinečnú špecifickosť. Naopak, sú schopní aktívne mobilizovať cez membránu buniek obličkových tubulov a črevného epitelu širokú škálu metabolitov, ako sú aminokyseliny, ióny, glukóza a osmolyty.
Funkcia tohto typu transportérov, ktorá bola širšie študovaná, je reabsorpcia glukózy prítomnej v moči.
Môže vám slúžiť: porovnávacia embryológia: Čo je to, aké štúdie, história, teórieTento reabsorpčný proces zahŕňa mobilizáciu uhľohydrátov z obličkových tubulov cez tubulárne epitelové bunky do lúmenu peritubulárnych kapilár. Byť izoformou s vysokou kapacitou a afinitou k glukóze SGLT-2, ktorý je hlavným daňovníkom.
Funkcia absorpcie glukózy v črevnom trakte sa pripisuje SGLT-1, dopravníkovi, ktorý napriek nízkej kapacite má vysokú afinitu k glukóze.
Tretí člen tejto rodiny, SGLT3, je exprimovaný v membránach buniek kostrových svalov a nervového systému, kde sa zdá, že nekonáva ako glukózový transportér, ale ako senzor koncentrácií tohto cukru v extracelulárnom médiu.
Funkcie izoforiem SGLT4, SGLT5 a SGLT6 doteraz neboli isté.
Odkazy
- Abramson J, Wright EM. Štruktúra a funkcia NA sympérerov s invertovanými opakovaniami. Curr Ace Struct Biol. 2009; 19: 425-432.
- Alvarado F, Crane RK. Štúdie mechanizmu črevnej absorpcie cukrov. Vii. Transport fenylglykozidu a jeho možný vzťah k inhibícii fhorizínu na prepravu SUG Actice SND SND MALÝ. Zákon o Biochim Biophys. 1964; 93: 116-135.
- Charron FM, Blanchard MG, Lapointe JY. Intracelulárna hypertonicita je zodpovedná za tok vody spojený s na_/glykóza Cotransport. Biophys J. 2006; 90: 3546-3554.
- Chen XZ, Coady MJ, LaPointe JY. Rýchle napätie svorkov. Biophys J. Devätnásť deväťdesiat šiestich; 71: 2544-2552.
- Dyer J, Wood Is, Palejwala A, Ellis A, Shirazi-Beechey SP. Expresia monosacharidových transportérov v čreve diabetických ľudí. AM J Physiol Gastrointest Liver Physiol. 2002; 282: G241-G248.
- Sátk M, Marks J, Unwin RJ. Predpokladané umiestnenie tkaniva a funkcia člena rodiny SLC5 SGLT3. Exp Physiol. 2017; 102 (1): 5-13.
- Turk E, Wright Em. MOMBRANE TOPOLOGY MOTIVS V RODINE SGG COTRANSporter. J memb biol. 1997; 159: 1-20.
- Turk E, Kim O, HE J, Whitelegge JP, Eskandari S, Lam JT, Kreman M, Zampihi G, Faull KF, Wright EM. Molekulárna charakterizácia Vibrio parahaemolyticus VSGLT: Model pre cukrovinky spojené s sodíkom. J Biol Chem. 2000; 275: 25711-25716.
- Taroni C, Jones S, Thornton JM. Analýza a predikcia väzbových miest uhľohydrátov. Proteín. 2000; 13: 89-98.
- Wright EM, Loo DD, Hirayama BA. Biológia transportérov glukózy sodného. Fyzikálny rev. 2011; 91 (2): 733-794.
- « Charakteristiky Magnetosféry Zeme, štruktúra, plyny
- Prvky magnetického poklesu a zemské magnetické pole »