

Charakteristiky jadrových pórov, funkcie, komponenty
- 4826
- 210
- Adrián Fajnor
On jadrová póra (Z gréckeho, póry = krok alebo tranzit) je „dvere“ jadra, ktoré umožňujú väčšinu transportu medzi nukleoplasmou a cytoplazmou. Jadrové póry sa spájajú s vnútornými a vonkajšími membránami jadra za tvorbu kanálov, ktoré slúžia na transport proteínov a RNA.
Slovo poro neodráža zložitosť príslušnej štruktúry. Z tohto dôvodu je vhodnejšie odkazovať na komplex jadrových pórov (CPN) namiesto jadrových pórov. CPN môže zažiť zmeny v jej štruktúre počas transportu alebo stavov bunkového cyklu.
Nedávno sa zistilo, že nukleoporíny, proteíny, ktoré tvoria CPN, majú dôležitú úlohu pri regulácii génovej expresie. Keď sa teda vyskytnú mutácie, ktoré ovplyvňujú funkciu nukleoporínov, patológie sa vyrábajú u ľudí, ako sú autoimunitné, kardiomyopatie, vírusové infekcie a rakovina.
[TOC]
Charakteristika
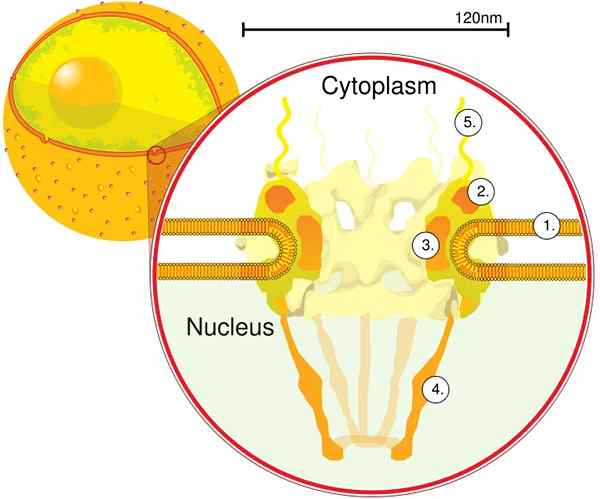
Prostredníctvom elektrónovej tomografie sa zistilo, že CPN má hrúbku 50 nm, vonkajší priemer medzi 80 a 120 nm a vnútorný priemer 40 nm. Veľké molekuly, ako je veľká podjednotka ribozómov (PM 1590 kDa), sa môžu vyvážať mimo jadra cez CPN. Odhaduje sa, že existuje medzi 2000 a 4000 CPN na jadro.
Molekulová hmotnosť jednotlivého CPN je približne medzi 120 a 125 mDa (1 mDa = 106 Da) v stavovcoch. Naopak, CPN je menší v kvasinkách, v ktorých má približne 60 mDa. Napriek obrovskej veľkosti CPN sú nukleoporíny vysoko zachované vo všetkých eukaryotoch.
Translokalizácia cez CPN je rýchly proces, ktorého rýchlosť je 1 000 translokácií/druhý. CPN však neurčuje smer samotného transportného toku.
Závisí to od gradientu RangTP, ktorý je väčší v jadre ako v cytosóle. Tento gradient je udržiavaná Ranom guanínovým výmenným faktorom.
Počas bunkového cyklu zažívajú CPN zostavy a cykly nezákonnosti. Zostava sa vyskytuje na rozhraní a bezprostredne po mitóze.
Funkcia
Kyselina ribonukleová (malá jadrová RNA, messengerová RNA, prenosová RNA), proteíny a ribonukleoproteíny (RNP) sa musia aktívne transportovať cez CPN. To znamená, že je potrebná energia hydrolýzy ATP a GTP. Každá molekula sa transportuje špecifickým spôsobom.
Všeobecne sú molekuly RNA plné komplexov RNP tvoriacich proteín, ktoré sa vyvážajú týmto spôsobom. Naopak, proteíny, ktoré sa aktívne transportujú do jadra, musia mať sekvenčný znak umiestnenia v jadre (SLN), majiteľ aminokyselinového odpadu s pozitívnym zaťažením (napríklad KKKrk).
Môže vám slúžiť: mesozómProteíny, ktoré sa exportujú do jadra.
Okrem uľahčovania transportu medzi jadrom a cytoplazmou sa CPN podieľajú na organizácii chromatínu, regulácii génovej expresie a opravou DNA. Nukleoporíny (Nups) Podporujú aktiváciu alebo represiu transkripcie, bez ohľadu na stav proliferácie buniek.
V kvasinkách sa NUPT nachádzajú v CNP jadrového obalu. V metazoári sú vo vnútri. Vykonávajú rovnaké funkcie vo všetkých eukaryotoch.
Dovoz látok
Prostredníctvom CPN je pasívne šírenie malých molekúl v oboch smeroch a aktívny transport, dovoz proteínov, vývoz RNA a ribonukleoproteínov (RNP) a obojsmerné molekuly raketle. Ten zahŕňa RNA, RNP a proteíny zapojené do signalizácie, biogenézy a náhrady.
Dovoz proteínov v jadre sa vyskytuje v dvoch krokoch: 1) väzba proteínu na cytoplazmatickú stranu CPN; 2) translokalizácia závislá od ATP cez CPN. Tento proces potrebuje hydrolýzu ATP a výmenu GTP/HDP medzi jadrom a cytoplazmou.
Podľa transportného modelu sa komplex prijímania proteínu pohybuje pozdĺž kanála cez Úniou, disociáciou a zjednotením opakovaných FG sekvencií nukleoporínov. Týmto spôsobom sa komplex presúva z jedného nukleoporínu do druhého vo vnútri CPN.
Vývoz látok
Je podobný ako v prípade dovozu. Run GTPase ukladá smerovanie prepravy cez CNP. RAN je molekulárny prepínač s dvoma konformačnými stavmi v závislosti od toho, či je spojený s HDP alebo GTP.
Dva špecifické regulačné proteíny spúšťajú konverziu medzi týmito dvoma stavmi: 1) cytosolický proteín aktivujúci GTPázu (GAP), ktorý produkuje hydrolýzu GTP, a tak prevádza RAN-GTP na RAN-GDP; a 2) výmenný faktor jadrovej guaniny (GEF), ktorý podporuje výmenu HDP pomocou GTP a konvertity RAN-GDP na RAN-GTP.
Cytosol obsahuje hlavne RAN-GDP. Jadro obsahuje hlavne RAN-GTP. Tento gradient týchto dvoch konformačných foriem RON prepravuje prepravu na príslušnej adrese.
Dovoz prijímača spolu s pozíciou je uľahčený Úniou k opakovaniu-fg. Ak sa dostanete na jadrovú stranu CNP, RAN-GTP sa pripojí k prijímači a uvoľní vašu pozíciu. RAN-GTP tak vytvára smer procesu importu.
Jadrový vývoz je podobný. RAN-GTP v jadre však podporuje spojenie pozície na vývozný príjemca. Keď sa vývozný prijímač pohybuje pórmi smerom k cytosólu, stretáva sa s gapom, ktorý indukuje hydrolýzu GTP na HDP. Nakoniec, prijímač je prepustený z jeho pozície a Ran-GDP v cytosole.
Môže vám slúžiť: polymorfonukleárne leukocytyPrenos RNA
Vývoz niektorých druhov RNA je podobný vývozu proteínov. Napríklad ARNT a RNAN (malé jadrové) používajú gradient RangTP a pomocou CPN sa prepravujú cez CPN. Vývoz zrelých ribozómov je tiež závislý od gradientu Rangtp.
MRNA sa vyváža veľmi odlišným spôsobom ako proteíny a iné RNA. Pre svoj vývoz mRNA tvorí komplex RNP Messenger (RNPM), v ktorom je molekula RNM obklopená stovkami proteínových molekúl. Tieto proteíny majú spracovanie, Uzatvorenie, Empalme a Polyadentilly z MNA.
Bunka musí byť schopná rozlišovať medzi RNPM s zrelým RNM a RNPM s nezrenou mRNA. MRNA, ktorá tvorí komplex RPNM, by mohla prijať topológie, ktoré je potrebné prepravovať na prepravu. Pred vstupom RNPM do CPN dochádza k kontrolnému kroku vykonávaným komplexmi Tramp a exozómovými proteínovými komplexmi.
Keď sa zostavuje zrelý RNPM, RPNM sa transportuje cez kanál pomocou transportného prijímača (NXF1-NXT1). Tento prijímač potrebuje hydrolýzu ATP (nie gradient RangTP) na stanovenie prestavby smerovania RNPM, ktorá dosiahne cytoplazmu.
Komplex jadrových pórov a kontrola génovej expresie
Niektoré štúdie naznačujú, že komponenty CPN by mohli ovplyvniť reguláciu genetickej expresie kontrolou chromatínovej štruktúry a jej prístupnosti k transkripčným faktorom.
V eukaryotoch novšieho vývoja je heterochromatín prednostne umiestnený na periférii jadra. Toto územie je prerušené euchromatínskymi kanálmi, ktoré sú udržiavané jadrovým košom CPN. Asociácia jadrového koša s euchromatínom súvisí s transkripciou génov.
Napríklad aktivácia transkripcie na úrovni CPN znamená interakciu komponentov jadrového koša s proteínmi, ako je histónová acetyltransferáza sága a vývozné faktory RNA.
Jadrový kôš je teda platformou pre mnohé gény údržby (Upratovanie) vysoko prepisované a gény silne vyvolané zmenami v podmienkach prostredia.
Komplex jadrových pórov a virologie
Infekcia vírusových buniek Eukaryot závisí od CPN. V každom prípade vírusovej infekcie závisí jej úspech od DNA, RNA alebo RPN, ktorý prechádza CPN, aby sa dosiahol jeho konečný cieľ, ktorým je replikácia vírusu.
Môže vám slúžiť: cytoplazmatické inklúzie: Čo sú, charakteristiky, funkcieVírus APE 40 (SV40) bol jedným z najviac študovaných modelov na skúmanie funkcie CPN v translokalizácii v jadre. Je to preto, že SV40 má malý genóm (5.000 báz).
Ukázalo sa, že transport vírusovej DNA je uľahčený proteínový proteín vírusu, ktoré chránia vírus, až kým jadro nedosiahne.
Komponenty
CPN je zabudovaný do jadrovej obálky a je tvorený približne 500 a 1 000 nupts. Tieto proteíny sú organizované v štrukturálnych subkomplexoch alebo moduloch, ktoré navzájom interagujú.
Prvým modulom je centrálna zložka alebo krúžok, vo vnútri póru v tvare piesočných hodín, ktorý je obmedzený ďalším krúžkom priemeru 120 nm na oboch stranách, intranukleárnym a cytoplazmatickým. Druhým modulom sú krúžky jadrových a cytoplazmy (každý z priemeru 120 nm) umiestnené okolo komponentu tvare Sandwate.
Tretím modulom je osem vlákien, ktoré sú vyčnievajúce z 120 nm kruhu v nukleoplazme a tvoria štruktúru tvare koša. Štvrtý modul sa skladá z vlákien, ktoré sa premietajú na bok cytoplazmy.
Komplex Y -the Y, pozostávajúci zo šiestich proteínov NUPS a SEH 1 a SEC 13, je najväčším a najlepšie charakterizovaným komplexom CNP. Tento komplex je nevyhnutnou jednotkou, ktorá je súčasťou lešenia CPN.
Napriek nízkej podobnosti medzi sekvenciami NUPS je lešenie CPN vo všetkých eukaryotoch vysoko zachované.
Odkazy
- Beck, m., Zranený, e. 2016. Komplex jadrových pórov: Pochopenie jeho funkcie prostredníctvom štrukturálneho pohľadu. Prírodné recenzie, biológia molekulárnych buniek, doi: 10.1038/nrm.2016.147.
- Ibarra, a., Hetzer, m.W. 2015. Proteíny jadrových pórov a kontrola funkcií genómu. Gény a vývoj, 29, 337-349.
- Kabachinski, G., Schwartz, T.Alebo. 2015. Komplex jadrových pórov - štruktúra a funkcia na prvý pohľad. Journal of Cell Science, 128, 423-429.
- Knocknhauer, k.A., Schwartz, T.Alebo. 2016. Komplex jadrových pórov ako flexibilná a dynamická brána. Cell, 164, 1162-1171.
- Ptak, c., Aitchison, J.D., Wozniak, r.W. 2014. Multifunkčný jadrový podľa komplexu: platforma na kontrolu génovej expresie. Súčasný názor na bunkovú biológiu, doi: 10.1016/j.CEB.2014.02.001.
- Stawicki, s.P., Steffen, J. 2017. Opublikácia: Komplex jadrových pórov - komplexný prehľad štruktúry a funkcie. International Journal of Academic Medicine, 3, 51-59.
- Tran, e.J., Wente, s.R. 2006. Dynamický jadrový podľa komplexov: Život na okraji. Cell, 125, 1041-1053.