

Fázy mitózy, charakteristiky, funkcie a organizmy
- 2886
- 20
- Alan Milota
Ten mitóza Je to proces delenia buniek, kde bunka produkuje geneticky identické dcérske bunky; Pre každú bunku sa generujú dve „dcéry“ s rovnakou chromozomálnou záťažou. Toto rozdelenie sa vykonáva v somatických bunkách eukaryotických organizmov.
Tento proces je jedným z štádií bunkového cyklu eukaryotických organizmov, ktorý sa chápe v 4 fázach: S (syntéza DNA), M (bunkové delenie), G1 a G2 (medziprodukty, kde sa vyskytujú mRNA a proteíny). Spoločne sa fázy G1, G2 a S považujú za rozhranie. Jadrové a cytoplazmatické delenie (mitóza a cytokinéza) tvoria poslednú fázu bunkového cyklu.
Prehľad mydózy. Zdroj: ViewAprabha [CC BY-SA 3.0 (https: // creativeCommons.Org/licencie/By-SA/3.0)] Na molekulárnej úrovni sa myitóza iniciuje aktiváciou kinázy (proteínu) nazývaného MPF (metučný faktor) a následnou fosforyláciou významného počtu zložiek buniek zložky. Ten umožňuje bunke prezentovať morfologické zmeny potrebné na vykonanie procesu delenia.
Mitóza je asexuálny proces, pretože progenitorová bunka a jej dcéry majú presne rovnaké genetické informácie. Tieto bunky sú známe ako diploid prenášaním úplného chromozomálneho zaťaženia (2N).
Meióza, na druhej strane, je proces delenia buniek, ktorý vedie k sexuálnej reprodukcii. V tomto procese diploidná kmeňová bunka replikuje svoje chromozómy a potom sa delí dvakrát v rade (bez replikácie jeho genetických informácií). Nakoniec sa generujú 4 dcérske bunky iba s polovicou chromozomálnej záťaže, ktoré sa nazývajú haploidy (N).
[TOC]
Všeobecnosť mitózy
Mitóza v jednobunkových organizmoch vo všeobecnosti produkuje dcérske bunky veľmi podobné jej rodičom. Naopak, počas vývoja mnohobunkových bytostí môže tento proces spôsobiť dve bunky s rôznymi charakteristikami (napriek tomu, že sú geneticky identické).
Táto bunková diferenciácia vedie k rôznym typom buniek, ktoré tvoria mnohobunkové organizmy.
Počas života organizmu sa bunkový cyklus vyskytuje nepretržite a neustále vytvára nové bunky, ktoré zase rastú a pripravujú sa na rozdelenie myitózou.
Bunkový rast a delenie sú regulované mechanizmami, ako je apoptóza (naprogramovaná bunková smrť), ktoré umožňujú udržiavanie rovnováhy a vyhýbajú sa nadmernému rastu tkanív. Týmto spôsobom je zabezpečené, že defektné bunky sa nahradia novými bunkami podľa požiadaviek a potrieb organizmu.
Aký význam má tento proces?
Schopnosť reprodukovať je jednou z najdôležitejších charakteristík všetkých organizmov (od jednobunkových až po mnohobunkové) a bunky, ktoré ho tvoria. Táto kvalita umožňuje zabezpečiť kontinuitu vašich genetických informácií.
Pochopenie procesov mitózy a meiózy malo základnú úlohu pri porozumení zaujímavých charakteristík buniek organizmov. Napríklad vlastnosť udržiavania počtu chromozómov z jednej bunky do druhej u jednotlivca a medzi jedincami toho istého druhu.
Keď v našej pokožke trpíme nejaký druh strihu alebo zranenia, pozorujeme, ako sa v priebehu dní vyžije poškodená pokožka. K tomu dochádza vďaka procesu myitosis.
Fázy a jeho vlastnosti
Všeobecne platí, že mytóza sleduje rovnakú procesnú sekvenciu (fázy) vo všetkých eukaryotických bunkách. V týchto fázach sa v bunke vyskytuje veľa morfologických zmien. Medzi nimi kondenzácia chromozómov, prasknutie jadrovej membrány, separácia bunky od extracelulárnej matrice a iných buniek a delenie cytoplazmy.
V niektorých prípadoch sa jadrové delenie a cytoplazmatické delenie považujú za rôzne fázy (mitóza a cytokinéza).
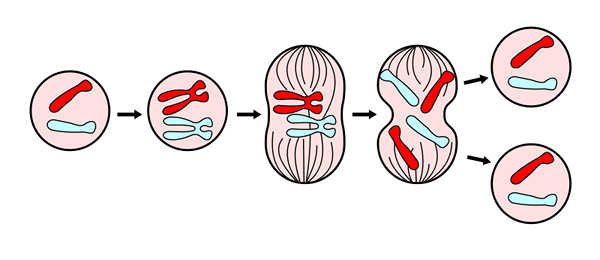
Pre lepšie štúdium a porozumenie procesu bolo určených šesť (6) fáz, nazývaných: Propáza, sľúbená, metafáza, anafáza a telofáza, potom sa zvažuje cytokinéza za šiestu fázu, ktorá sa začína vyvíjať počas anafázy.
 Telofáza je poslednou fázou mitózy. Prevzaté z https: // commons.Wikimedia.Org/wiki/súbor: Mitosepanel.Jpg. Cez Wikimedia Commons
Telofáza je poslednou fázou mitózy. Prevzaté z https: // commons.Wikimedia.Org/wiki/súbor: Mitosepanel.Jpg. Cez Wikimedia Commons Tieto fázy sa študovali už od devätnásteho storočia ľahkým mikroskopom, takže dnes sú ľahko rozpoznateľné podľa morfologických charakteristík bunky, ako je chromozomálna kondenzácia a tvorba mitotického vretena.
Prorok
 Prorok. Leomonaci98 [CC BY-SA 4.0 (https: // creativeCommons.Org/licencie/By-SA/4.0)], z Wikimedia Commons
Prorok. Leomonaci98 [CC BY-SA 4.0 (https: // creativeCommons.Org/licencie/By-SA/4.0)], z Wikimedia Commons Profase je prvý viditeľný prejav delenia buniek. V tejto fáze môžete vidieť vzhľad chromozómov ako rozlíšiteľné formy v dôsledku progresívneho zhutnenia chromatínu. Táto kondenzácia chromozómov sa začína fosforyláciou molekúl h1 H1 pomocou MPF kinázy.
Kondenzačný proces pozostáva z kontrakcie, a teda k zníženiu veľkosti chromozómov. K tomu dochádza v dôsledku varenia chromatínových vlákien, ktoré vytvárajú ľahšie vytlačené štruktúry (mitotické chromozómy).
Chromozómy, ktoré boli predtým duplikované počas obdobia bunkového cyklu, získavajú vzhľad dvojitého vlákna nazývaného sesterské chromatidy, tieto vlákna zostávajú zjednotené prostredníctvom regiónu nazývaného Centromero. V tejto fáze tiež jadrá zmiznú.
Môže vám slúžiť: Cromafin bunky: charakteristiky, histológia, funkcieTvorba mitotických vretena
Autor: Silvia3 [GFDL (http: // www.GNU.Org/copyleft/fdl.HTML) alebo CC By-SA 4.0 (https: // creativeCommons.Org/licencie/By-SA/4.0)], z Wikimedia Commons Počas prorázy sa tvorí mitotické vreteno, ktoré pozostáva z mikrotubulov a proteínov, ktoré tvoria sadu vlákien.
Keď sa vytvára vreteno, mikrotubuly cytoskeletu (deaktiváciou proteínov, ktoré si udržiavajú ich štruktúru), sú zostavené, čím sa poskytuje potrebný materiál na tvorbu uvedeného mitotického vretena.
Centrá (organela bez membrány, funkčné v bunkovom cykle), duplikát v rozhraní, pôsobí ako montážna jednotka mikrotubulov vretena. V živočíšnych bunkách má centrum v strede, pár centriolov; Ale vo väčšine rastlinných buniek chýbajú.
Duplicitné centrá, začínajú oddeliť od druhého, zatiaľ čo v každom z nich sú zostavené mikrotubuly vretena, začínajú migrovať na opačné konce bunky.
Na konci prorázy sa začína prasknutie jadrového zabalenia, vyskytujúce sa v samostatných procesoch: Útok MAS jadrového póru, jadrový list a jadrové membrány. Táto prestávka umožňuje mitotické vreteno a chromozómy začínajú interagovať.
Sľubovať
Leomonaci98 [CC BY-SA 4.0 (https: // creativeCommons.Org/licencie/By-SA/4.0)] V tomto štádiu bola jadrová obálka úplne fragmentovaná, takže mikrotubuly vretena napadnú do tejto oblasti, interagujúca s chromozómami. Obidve centrá sa oddelili, každé v póloch mitotického vretena v opačných extrémoch buniek.
Teraz mitotické vreteno zahŕňa mikrotubuly (ktoré sa rozširujú od každého centra do stredu bunky), centrá a pár osterov (štruktúry s radiálnym rozložením krátkych mikrotubulov, ktoré sú nasadené z každého centra).
Každý z chromatidov sa vyvinul, špecializovaná proteínová štruktúra nazývaná Cinetocoro, ktorá sa nachádza v centromere. Tieto knetocory sú umiestnené v opačných smeroch a niektoré mikrotubuly sú priľnavé, nazývané mikrotubuly cinnetocoro.
Tieto mikrotubuly pripojené k Cinetocoro začínajú pohybovať chromozóm od konca, ktorý sa rozširujú; Niektorí z pólu a iné z opačného pólu. To vytvára efekt „ťahania a zmenšenia“, ktorý pri stabilizácii umožňuje chromozóm končiť medzi koncami bunky.
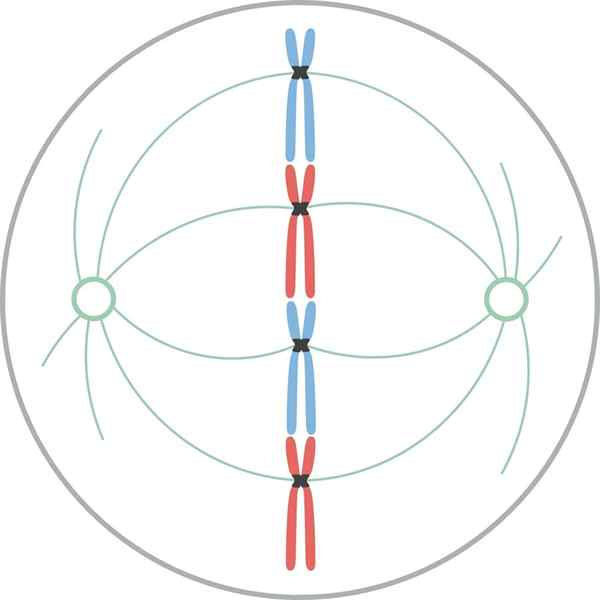
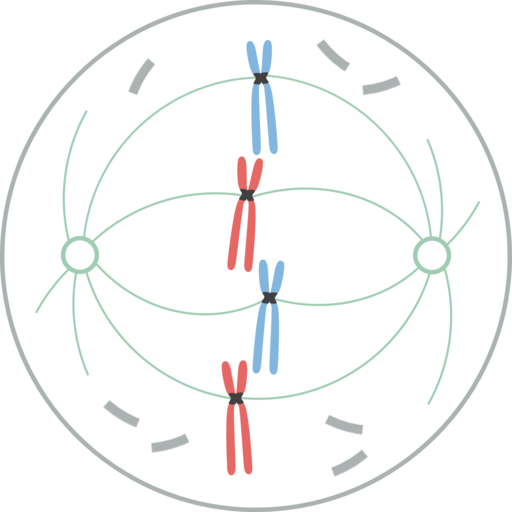
Metafáza
Chromozómy zarovnané v rovníkovej doštičke bunky počas mýtickej metafázy V metafáze sú centrá umiestnené na opačných koncoch buniek. Vreteno ukazuje jasnú štruktúru, v ktorej strede sa nachádzajú chromozómy. Centroméry týchto chromozómov sú fixované na vlákna a zarovnané v imaginárnej rovine nazývanej metafázická plak.
Chromatidové cipnetocory zostávajú pripevnené k mikrotubulám Cinetocoro. Mikrotubuly, ktoré nedodržiavajú Knetocoros a siahajú z opačných pólov vretena, teraz vzájomne interagujú. V tomto bode sú mikrotubuly z osterov v kontakte s plazmatickou membránou.
Tento rast a interakcia mikrotubulov dokončuje štruktúru mitotického vretena a dáva vzhľad „vtáčej klietky“.
Morfologicky je táto fáza fáza, ktorá sa javí menšie zmeny, takže sa považovala za fázu pokoja. Aj keď nie sú ľahko značné, v ňom sa vyskytuje veľa dôležitých procesov, okrem toho, že je najdlhším štádiom mitózy.
Anafáza
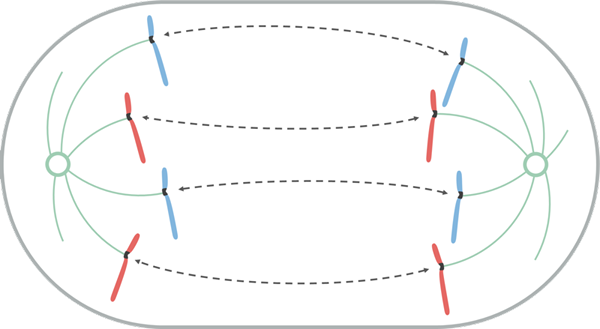 Zdroj: Leomonaci98 [CC BY-SA 4.0 (https: // creativeCommons.Org/licencie/By-SA/4.0)], z Wikimedia Commons
Zdroj: Leomonaci98 [CC BY-SA 4.0 (https: // creativeCommons.Org/licencie/By-SA/4.0)], z Wikimedia Commons Počas anafázy sa každý pár chromatidov začína oddeliť (inaktiváciou proteínov, ktoré ich udržiavajú pohromade). Oddelené chromozómy sa pohybujú smerom k opačným koncom bunky.
Tento pohyb migrácie je spôsobený skutočnosťou, že mikrotubuly Catocoro de Acortan, ktorý vytvára efekt „ťahania“, ktorý spôsobuje, že každý chromozóm sa pohybuje od svojho centromero. V závislosti od umiestnenia centroméry v chromozóme môže mať počas jeho posunu konkrétnu formu, ako je V alebo j.
Mikrotubuly, ktoré nie sú priľnavé k Knetocoro, rastú a predlžujú sa tubulínovou adhéziou (proteín) a pôsobením motorických proteínov, ktoré sa k nim pohybujú, čo umožňuje kontakt medzi nimi zastaviť. Keď sa pohybujú od seba, robia to aj vretenové stĺpy a predlžujú bunku.
Na konci tejto fázy sú skupiny chromozómov umiestnené na opačných koncoch mitotického vretena, takže každý koniec bunky je s úplnou a ekvivalentnou sadou chromozómov.
Telofáza
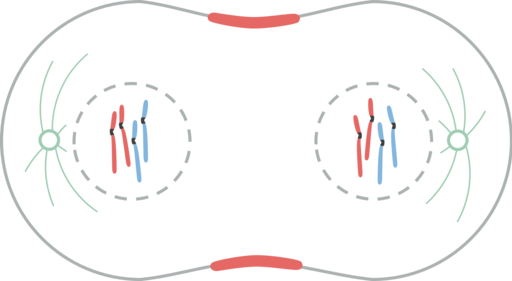 Telofáza. Leomonaci98 [CC BY-SA 4.0 (https: // creativeCommons.Org/licencie/By-SA/4.0)]
Telofáza. Leomonaci98 [CC BY-SA 4.0 (https: // creativeCommons.Org/licencie/By-SA/4.0)] Telofáza je poslednou fázou jadrového delenia. Mikrotubuly Cinetocoro sa rozpadajú, zatiaľ čo polárne mikrotubuly sa ďalej predlžujú.
Jadrová membrána sa začína tvoriť okolo každej hry chromozómov pomocou jadrových zábalov materskej bunky, ktoré boli ako cytoplazma vezikúl.
V tomto štádiu sú chromozómy, ktoré sú v bunkových póloch, úplne odrádzajú v dôsledku defosforylácie histónových molekúl (H1). Tvorba prvkov jadrovej membrány je riadená niekoľkými mechanizmami.
Počas anafázy sa v proráze začalo deforilovať mnoho fosforylovaných proteínov. To umožňuje začiatok telofázy, jadrové vezikuly sa začnú znovu zostaviť, spojené s povrchom chromozómov.
Môže vám slúžiť: Chondrocyty: Charakteristiky, histológia, funkcie, kultiváciaNa druhej strane, jadrová póra je opätovne assembla, ktorá umožňuje čerpanie jadrových proteínov. Proteíny jadrovej laminy sú defosforylované, čo im umožňuje znova sa spájať, dokončiť tvorbu uvedenej jadrovej laminy.
Nakoniec, keď sú chromozómy úplne odrádzané, sa syntéza RNA reštartuje, čím sa jadro opäť tvorí, a tým dokončí tvorbu nových jadier rozhrania dcérskych buniek.
Cytokinéza

Cytocinesis sa považuje za udalosť oddelenú od jadrového delenia a bežne v typických bunkách sprevádza proces cytoplazmatického delenia každú myitózu, začínajúc anafázou. Niekoľko štúdií ukázalo, že v niektorých embryách sa pred cytoplazmatickým delením vyskytujú viac jadrových divízií.
Tento proces začína vzhľadom drážky alebo rozštepu, ktorá je označená v rovine metafázického plaku, čím sa zabezpečí, že k rozdeleniu dôjde medzi skupinami chromozómov. Miesto rozštiepenia je označené konkrétne mitotickým vretenom, mikrotubuly ásterov.
V označenej štrbine je séria mikrofilamentov tvoriacich krúžok nasmerovaný na cytoplazmatickú stranu bunkovej membrány, z veľkej časti zloženej z aktínu a myozínu. Tieto proteíny navzájom interagujú, čo umožňuje kontrakciu kruhu okolo drážky.
Táto kontrakcia je generovaná posuvou vlákien týchto proteínov, keď sa navzájom interagujú, rovnakým spôsobom, ako to robia napríklad vo svalových tkanivách.
Kontrakcia kruhu sa prehĺbi tým, že vyvíja efekt „štipka“, ktorý nakoniec rozdeľuje rodičovskú bunku, čo umožňuje separáciu dcérskych buniek, s jeho cytoplazmatickým obsahom vo vývoji.
Cytocinéza v rastlinných bunkách
Rastlinné bunky majú bunkovú stenu, takže ich cytoplazmatický proces delenia sa líši od procesu opísaného vyššie a začína v telofáze.
Tvorba novej bunkovej steny začína zostavovať mikrotubuly zvyškového vretena, čím predstavuje framoplast. Táto valcová štruktúra je tvorená dvoma mikrotubulami, ktoré sa spájajú na svojich koncoch a ktorých pozitívne póly sú zabudované do elektronického plaku v rovníkovej rovine.
Malé vezikuly z Golgiho prístroja, plné prekurzorov bunkovej steny, sa pohybujú cez mikrotubuly framoplastu do rovníkovej oblasti, ktoré sa kombinujú za vzniku bunkového plaku. Obsah vezikúl sa na tejto doske oddeľuje, keď rastie.
Táto doska rastie a spája sa s plazmovou membránou pozdĺž obvodu buniek. K tomu dochádza v dôsledku konštantnej reorganizácie mikrotubulov framoplastu na okraji dosky, čo umožňuje viac vezikúl pohybovať sa smerom k tejto rovine a vyprázdniť jej obsah.
Týmto spôsobom dochádza k cytoplazmatickej separácii dcérskych buniek. Nakoniec obsah bunkovej dosky vedľa celulózových mikrovlákanov vo vnútri umožňuje dokončiť tvorbu novej bunkovej steny.
Funkcia
Mitóza je mechanizmus delenia v bunkách a je súčasťou jednej z fáz bunkového cyklu v eukaryotoch. Jednoduchým spôsobom môžeme povedať, že hlavnou funkciou tohto procesu je reprodukcia bunky v dvoch dcérskych bunkách.
V prípade jednobunkových organizmov je delenie buniek generovanie nových jedincov, zatiaľ čo pre mnohobunkové organizmy je tento proces súčasťou rastu a správneho fungovania úplného tela (bunkové delenie vytvára vývoj tkanív a udržiavanie štruktúr).
Proces mytózy je aktivovaný podľa požiadaviek organizmu. Napríklad u cicavcov začnú červené krvinky (erytrocyty) rozdeliť viac buniek, keď telo potrebuje lepšie zachytenie kyslíka. Podobne sa reprodukujú biele krvinky (leukocyty), keď je potrebné bojovať proti infekcii.
Naopak, niektoré špecializované živočíšne bunky, prakticky chýbajú proces mitózy alebo je veľmi pomalý. Príkladom sú nervové bunky a svalové bunky).
Všeobecne sú to bunky, ktoré sú súčasťou spojivového a štrukturálneho tkaniva organizmu a ktorého reprodukcia je potrebná iba vtedy, keď má bunka určitú defekt alebo zhoršenie a je potrebné ju nahradiť.
Regulácia rastu a delenie buniek.
Bunkový rast a systém delenia buniek sú v mnohobunkových organizmoch oveľa zložitejšie ako v jednobunkových. V druhom prípade je reprodukcia v podstate obmedzená dostupnosťou zdrojov.
V živočíšnych bunkách je delenie zadržané, až kým neexistuje pozitívny signál, ktorý tento proces aktivuje. Táto aktivácia prichádza vo forme chemických signálov zo susedných buniek. To umožňuje zabrániť neobmedzenému rastu tkanív a reprodukcii defektných buniek, ktoré môžu vážne poškodiť životnosť organizmu.
Môže vám slúžiť: bazálna membrána: Charakteristiky, štruktúra a funkcieJedným z mechanizmov, ktoré kontrolujú násobenie buniek, je apoptóza, kde bunka zomrie (v dôsledku produkcie určitých proteínov, ktoré aktivujú sebazničenie), ak má značné poškodenie alebo je infikované vírusom.
Existuje tiež regulácia vývoja buniek prostredníctvom inhibície rastových faktorov (ako je proteín). Bunky teda zostávajú v rozhraní, bez toho, aby postupovali do m fázy bunkového cyklu.
Organizmy, ktoré to vykonávajú
Proces mitózy sa vykonáva v drvivej väčšine eukaryotických buniek, z jednobunkových organizmov, ako sú kvasinky, ktoré ho používajú ako asexuálny proces reprodukcie, na komplexné multicelulárne organizmy, ako sú rastliny a zvieratá a zvieratá.
Aj keď vo všeobecnosti je bunkový cyklus rovnaký pre všetky eukaryotické bunky, existujú pozoruhodné rozdiely medzi jednobunkovými a mnohobunkovými organizmami. V prvom rade je rast a delenie buniek uprednostňovaný prírodným výberom. V mnohobunkových organizmoch je proliferácia obmedzená prísnymi kontrolnými mechanizmami.
V reprodukcii jednobunkových organizmov sa zrýchľuje, pretože bunkový cyklus funguje neustále a dcérske bunky sa rýchlo púšťajú do myitózy, aby pokračovali v tomto cykle. Zatiaľ čo bunky mnohobunkových organizmov trvá oveľa viac času na rast a rozdelenie.
Existujú tiež určité rozdiely medzi mitotickými procesmi rastlinných a živočíšnych buniek, ako v niektorých fázach tohto procesu, v zásade však mechanizmus funguje podobne v týchto organizmoch.
Bunkové delenie v prokaryotických bunkách
Prokaryotická bunka Vo všeobecnosti rastú prokaryotické bunky a sú rozdelené rýchlejšie ako eukaryotické bunky.
Organizmy s prokaryotickými bunkami (zvyčajne jednobunkovými alebo v niektorých mnohobunkových prípadoch) nemajú jadrovú membránu, ktorá izoluje genetický materiál vo vnútri jadra, takže je dispergovaný v bunke, v oblasti nazývanej nukleoid. Tieto bunky majú hlavný kruhový chromozóm.
Bunkové delenie v týchto organizmoch je potom oveľa priamejšie ako v eukaryotických bunkách, chýba mu opísaný mechanizmus (mitóza). V nich sa reprodukcia vykonáva procesom nazývaným binárne štiepenie, kde replikácia DNA začína v špecifickom mieste kruhového chromozómu (pôvod replikácie alebo orika).
Vytvárajú sa dva pôvody, ktoré migrujú na opačné strany bunky, keď dôjde k replikácii, a bunka sa tiahne, až kým nedosiahne dvojnásobok veľkosti. Na konci replikácie rastie bunková membrána do cytoplazmy a rozdeľuje progenitorovú bunku na dve dcéry s rovnakým genetickým materiálom.
Vývoj mitózy
Vývoj eukaryotických buniek so sebou priniesol zvýšenie zložitosti genómu. Z toho vyplývalo rozvoj prepracovanejších mechanizmov rozdelenia.
Čo predchádzalo mitóze?
Existujú hypotézy, ktoré navrhujú, že bakteriálne delenie je predchodcom mechanizmu mitózy. Zistil sa určitý vzťah medzi proteínmi spojenými s binárnym štiepením (ktoré môžu byť tie, ktoré ukotvujú chromozómy na špecifické miesta plazmatickej membrány dcér) s tubulínom a aktínom eukaryotických buniek.
Niektoré štúdie naznačujú určité zvláštnosti v rozdelení moderných jednobunkových protistov. V nich jadrová membrána zostáva počas mitózy nedotknutá. Replikované chromozómy zostávajú ukotvené na určitých miestach tejto membrány a oddeľujú sa, keď sa jadro začne rozširovať počas delenia buniek.
To ukazuje určitú zhodu s procesom binárneho štiepenia, kde sú replikované chromozómy fixované na určité miesta v bunkovej membráne. Hypotéza potom navrhuje, aby protisti, ktorí predstavujú túto kvalitu počas svojho bunkového delenia, mohli zachovať túto charakteristiku predkových bunkových buniek.
V súčasnosti, vysvetlenia, prečo ešte neboli vyvinuté eukaryotické bunky mnohobunkových organizmov, je potrebné, aby sa jadrová membrána rozpadla počas procesu delenia buniek.
Odkazy
- Albarracín, a., & Telulón,. Do. (1993). Teória buniek v devätnástom storočí. Akal vydania.
- Alberts, b., Johnson, a., Lewis, J., Raff, m., Roberth, K., & Walter, P. (2008). Biológia molekulárnej bunky. Garland Science, Taylor a Francis Group.
- Campbell, n., & Reece, J. (2005). Biológia 7th Vydanie, AP.
- Griffiths, a. J., Lewontin, r. C., Miller, J. H., & Suzuki, D. Tón. (1992). Úvod do genetickej analýzy. McGraw-Hill Inter-American.
- Karp, G. (2009). Bunková a molekulárna biológia: koncepty a experimenty. John Wiley & Sons.
- Ubytovňa, h., Darnell, J. A., Berk, a., Kaiser, C. Do., Krieger, m., Scott, m. P., & Matsudaira, P. (2008). Biológia mäkkýšov. Macmillan.
- Segura-Valdez, m. D. L., Cruz-Gómez, s. D. J., López-Cruz, R., Zavala, G., & Jiménez-García, L. F. (2008). Vizualizácia mitózy pomocou mikroskopu atómovej sily. Tip. Špecializovaný časopis na chemické biologické vedy, 11 (2), 87-90.