

Štruktúra cytozínu, funkcie, vlastnosti, syntéza
- 910
- 85
- MUDr. Žigmund Boška
Ten Cytozín Je to báza dusíka pyrimidínu, ktorá slúži na biosyntézu cytidínu-5'-monofosfátu a dexicitidín-5'-monofosfátu. Tieto zlúčeniny slúžia na biosyntézu, respektíve z kyseliny deoxyribonukleovej (DNA) a kyseliny ribonukleovej (RNA). DNA ukladá genetické informácie a RNA má rôzne funkcie.
U živých bytostí sa cytozín nenachádza voľne, ale bežne ribonukleotidy alebo deoxyribonukleotidy. Oba typy zlúčenín majú fosfátovú skupinu, ribózu a dusíkovú bázu.
Zdroj: Vesprcom [verejná doména] Uhlík 2 ribózy má oxhydrillo (-OH) skupinu v ribonukleotidoch a atóm vodíka (-H) v deoxyribonukleotidoch. V závislosti od počtu prítomných fosfátových skupín je Citidín-5'-monofosfát (CMP), Citidín-5'-difosfát (CDP) a Citidín-5'-triffosfát (CTP) (CTP) (CTP).
Desoxygenované ekvivalenty sa nazývajú desoxicitidín-5'-monofosfát (DCMP), desoxicitidín-5'-difosfát (DCDP) a desoxicitidín-5'-triffosfát (DCTP).
Cytozín sa vo svojich rôznych formách podieľa na rôznych funkciách, ako je biosyntéza DNA a RNA, biosyntéza glykoproteínov a regulácia expresie génu.
[TOC]
Štruktúra
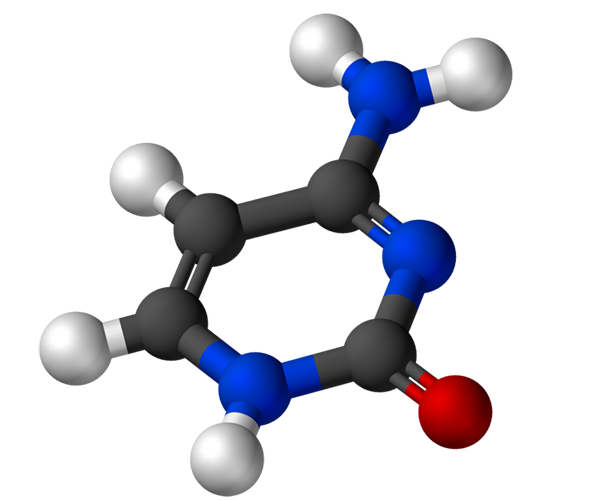
Cytozín, 4-amino-2-hydroxipirimidín, má empirický vzorec C4H5N3Alebo, ktorého molekulová hmotnosť je 111,10 g/mol a je purifikovaná ako biely prášok.
Štruktúra cytozínu je heterocyklický, aromatický a planárny kruh. Maximálna vlnová dĺžka absorbancie (ʎMaximálny) Je 260 nm. Teplota fúzie cytozínu presahuje 300 ° C.
Na vytvorenie nukleotidu sa cytozín kovalentne viaže prostredníctvom dusíka 1. Uhlík 5 'je esterifikovaný fosfátovou skupinou.
Biosyntéza
Biosyntéza pyrimidínu nukleotidov má spoločnú cestu, ktorá pozostáva zo šiestich krokov katalyzovaných enzýmami. Cesta začína biosyntézou karbamil-fosfátu. V prokaryotoch je iba jeden enzým: syntáza fosforečnanu karbamilu. Toto je zodpovedné za syntézu pyrimidínov a glutamínu. V eukaryotoch existujú karbamil fosfáty syntázy I a II, ktoré sú zodpovedné za biosyntézu glutamínu a pyrimidínu.
Druhý krok pozostáva z tvorby N-CarbamoAspartato, z karboilového fosfátu a aspartátu, reakcie katalyzovanej transpartátom transcabamoilou (ATCASA).
Tretím krokom je syntéza l-dihydrotoátu, ktorý vytvára uzavretie pyrimidínového kruhu. Tento krok je katalyzovaný dihydroottázou.
Štvrtým krokom je tvorba Orotato, ktorá je redoxnou reakciou katalyzovanou dihydroorotátovou dehydrogenázou.
Piaty krok pozostáva z tvorby goldylátu (OMP) s použitím fosforibozylpyrofosfátu (PRPP) ako substrátu a fosforibozyl -orotátovej transferázy ako katalyzátora ako katalyzátora ako katalyzátora ako katalyzátor.
Šiesty krok je tvorba Uridilaato (uridin-5'-monofosfát, UMP), reakcia katalyzovaná OMP-decarboxylázou.
Môže vám slúžiť: Flora a fauna z HidalgoNasledujúce kroky pozostávajú z fosforylácie UMP, katalyzovanej kinázami, za vzniku UTP a prenosu aminoskupiny z glutamínu do UTP za vzniku CTP, reakcia katalyzovaná CTP syntetázou.
Regulácia biosyntézy
U cicavcov sa regulácia uskutočňuje pri syntáze fosfátovej fosfátovej karbamilu.
Carbamoil SyntasA II je regulovaná negatívnou spätnou väzbou. Jeho regulátory, UTP a PRPP, sú inhibítorom a aktivátorom tohto enzýmu.
V tkanivách, ktoré nie sú pečeňové, je karbamil fosfát Syntáza II je jediným zdrojom fosforečnanu karbamilu fosforečnanu. Zatiaľ čo v pečeni, v podmienkach nadmerného amoniaku, produkuje syntáza fosfátu karbamilu I, v mitochondriách fosforečnan, ktorý sa transportuje do cytosólu, odkiaľ vstupuje do cesty biosyntézy pyrimidínov.
Ďalším regulačným bodom je OMP-decarboxyláza, ktorá je regulovaná konkurenčnou inhibíciou. Produkt jeho reakcie, UMP, súťaží s OMP o väzbové miesto v OMP-Descarboxilasa.
Pyrimidíny, podobne ako cytozín, sa recyklujú
Recyklácia pyrimidínov má ako funkciu opätovné použitie pyrimidínov bez potreby biosyntézy Novo a vyhnúť sa degradatívnej ceste. Recyklačná reakcia je katalyzovaná fosforibosyltransferázou pyrimimidínom. Všeobecná reakcia je nasledovná:
Pyrimidín + prpp -> nukleozid pyrimidín 5 '-monofosfát + PPI
U stavovcov sa fosforibosyltransferáza pyrimimidín nachádza v erytrocytoch. Pyrimidíny substrát tohto enzýmu sú Uracil, Timina a Orotato. Cytozín sa nepriamo recykluje z uridínu-5'-monofosfátu.
Funkcia v biosyntéze DNA
Počas replikácie DNA sa informácie obsiahnuté v DNA skopírujú v DNA prostredníctvom DNA polymerázy.
Biosyntéza RNA potrebuje deoxynukleotidy Tryfosfát (DNTP), konkrétne: dexitimid tryposfát (DTTP), dexicitidy triffosfát (DCTP), de -alxyxyadenín Tryphyse (DATP) a dexiguanín Tryfosfát (DGTP). Reakcia je:
(DNA)n odpad + Dntp -> (dna)N+1 odpad + PPI
Hydrolýza anorganického pyrofosforečnanu (PPI) poskytuje energiu na biosyntézu RNA.
Funkcia pri stabilizácii štruktúry DNA
V dvojitej špirále DNU je purín, reťazca, spojený s pyrimidínom opačného reťazca, vodíkovými väzbami. Cytozín je teda vždy spojený s guanínom tromi vodíkovými väzbami: adenín je spojený s timinom dvoma vodíkovými väzbami.
Môže vám slúžiť: Dôležitosť biológieVodíkové väzby sú prerušené, keď je čistý natívny roztok DNA pri pH 7 vystavený teplotám vyšším ako 80 ° C. To spôsobí, že dvojitá vrtuľa DNA vytvorí dva samostatné reťazce. Tento proces je známy ako denaturácia.
Teplota, pri ktorej je denaturovaná 50% DNA, je známa ako teplota topenia (TM). Molekuly DNA, ktorých guaníny a cytozín.
Vyššie uvedené je experimentálny test, že väčší počet vodíkových väzieb lepšie stabilizuje natívne molekuly DNA.
Funkcia bohatých oblastí v cytozíne v DNA
Nedávno sa zistilo, že DNA jadra ľudských buniek môže prijať interkalované motívové štruktúry (IM). Tieto štruktúry sa vyrábajú v cytozínových oblastiach.
Štruktúra IM pozostáva zo štyroch reťazcov DNA, na rozdiel od klasickej DNA s dvojitou vrtuľou, ktorá má dva reťazce. Presnejšie povedané, dva paralelné duplexné reťazce sú rozptýlené v orientácii antiparallela a sú udržiavané spolu pomocou pár hemiprotonovaných cytozínov (C: C: C: C: C: C: C: C: C:+).
V ľudskom genóme sa IM štruktúry nachádzajú v regiónoch, ako sú promótory a teloméry. Počet štruktúr IM je vyšší počas fázy G1/S bunkového cyklu, v ktorom je transkripcia vysoká. Tieto oblasti sú miesta rozpoznávania proteínov zapojené do aktivácie transkripčných strojov.
Na druhej strane, v regiónoch bohatých na guanínové základne (c) po sebe idúce DNA má tendenciu adoptovať vrtuľový tvar A v dehydratačných podmienkach. Táto forma je typická pre RNA a dvojité zmiešané pásy DNA-ARN počas transkripcie a replikácie a v určitých časoch, keď je DNA spojená s proteínom.
Ukázalo sa, že regióny s po sebe idúcimi cytozínovými základňami vytvárajú elektropozitívnu náplasť v hlavnej štrbine DNA. Preto sa predpokladá, že tieto oblasti sa viažu na proteíny, ktoré predisponujú určité genomické oblasti genetickej krehkosti.
Funkcia v biosyntéze RNA
Počas prepisu sa informácie obsiahnuté v DNA skopírujú v RNA cez Pon polymerázovú RNA. Potreby biosyntézy RNA Tryppost Nukleozidy (NTP), menovite: Tryphyse citidín (CTP), Tryposfát urridín (UTP), adenín Tryposfát (ATP) a Guanín trypfosfátu (GTP). Reakcia je:
Môže vám slúžiť: Flora a fauna z Campeche: Reprezentatívne druhy(RNA)n odpad + NTP -> (RNA)N+1 odpad + PPI
Hydrolýza anorganického pyrofosforečnanu (PPI) poskytuje energiu na biosyntézu RNA.
Funkcia v biosyntéze glykoproteínov
Postupný prenos hexóz na vytvorenie oligosacharidov, O-ligátov na proteíny, sa uskutoční z nukleotidových prekurzorov.
U stavovcov je posledný krok biosyntézy O ligosacharidov oligosacharidov pri pridávaní dvoch odpadu z kyseliny sinálneho kyseliny (N-acetylneuramín) z prekurzoru citidínu-5'-monofosfátu (CMP) (CMP). Táto reakcia sa vyrába v trans vrece Golgi.
Liečba cytozínu a chemoterapie proti rakovine
Kyselina tetrahydrofolát (FH4) je zdroj skupiny -ch3, A je to potrebné pre biosyntézu DTMP z skládky. Vytvára sa aj FH2. Redukcia FH2 na FH4 vyžaduje folát a NADPH reduktázu. Pri liečbe rakoviny sa používajú niektoré inhibítory reduktázy listov.
Metotrexan je konkurenčný inhibítor. Folátová reduktáza sa spája s 100 -krát väčšou afinitou k tomuto inhibítorovi ako k jeho substrátu. Aminapterín pôsobí podobne.
Inhibícia folátovej reduktázy nepriamo bráni biosyntéze DTMP, a teda aj indikácia DCTP. Priama inhibícia dochádza prostredníctvom inhibítorov Simidile Syntetázy, ktorá katalyzuje DTMP z skládky. Tieto inhibítory sú 5-fluóruracil a 5-fluór-2-dioxiuridín.
Napríklad 5-fluóracil nie je inhibítorom, ale najprv sa stáva recyklačnou cestou, v deoxyuridovom mofosfáte D (Fdump), ktorý sa viaže na syntetizáciu timilátu a inhibuje ju.
Látky analogické glutamínu, azaserínu a acivicínu, inhibujú amidotransferázu glutamín. Azarine bola jednou z prvých objavených látok, ktoré pôsobia ako samovražedná inaktivácia.
Odkazy
- Assi, h.Do., Garaví, m., González, C., A damha, m.J. 2018. I-motif DNA: Štrukturálne vlastnosti a význam pre bunkovú biológiu. Výskum jadrových kyselín, 46: 8038-8056.
- Bohinski, r. 1991. Biochémia. Addison-Wesley Iberoamericana, Wilmington, Delaware.
- Devlin, T.M. 2000. Biochémia. Redakčné Reverté, Barcelona.
- Ubytovňa, h., Berk, a., Zipurski, s.L., Matsudaria, P., Baltimore, D., Darnell, J. 2003. Bunková a molekulárna biológia. Redakcia Medica Panamericana, Buenos Aires, Bogota, Caracas, Madrid, Mexiko, Sāo Paulo.
- Nelson, D. L., Cox, m. M. 2008. Lehninger-Principles of Biochemistry. W.H. Freeman, New York.
- Voet, D. a Voet, J. 2004. Biochémia. John Wiley a Sons, USA.