

Štruktúra muskarínových receptorov, typy a ich funkcie, antagonisty
- 1441
- 202
- Valentín Dula
Ten muškarinické receptory Sú to molekuly, ktoré sprostredkujú acetylcholín (ACh) a sú umiestnené v postsynaptickej membráne synapsií, v ktorých je uvoľňovaný uvedený neurotransmiter; Jeho meno pochádza z jeho citlivosti na muskarínový alkaloid, ktorý produkuje huba Amanita Muscaria.
V centrálnom nervovom systéme je niekoľko neurónových súprav, ktorých axóny uvoľňujú acetylcholín. Niektorí z nich končia v samotnom mozgu, zatiaľ čo väčšina tvoria motorické dráhy pre kostrové svaly alebo efektorové dráhy autonómneho nervového systému pre žľazy a srdce a hladké svaly.
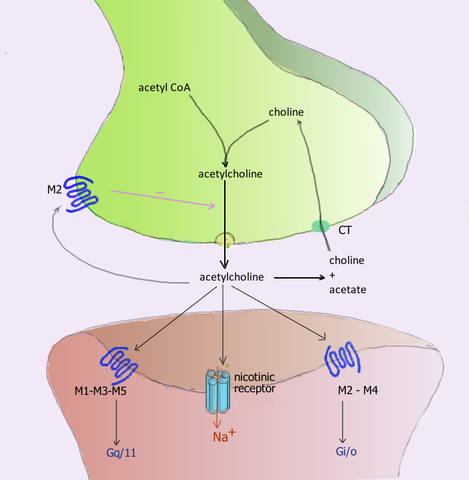
Neuroreceptor acetylcholínu počas synapsy a ich príslušné receptory v postsynaptickej membráne (zdroj: používateľ: pancrat [GFDL 1.2 (http: // www.GNU.Org/licencie/staré licenky/FDL-1.2.html)] Via Wikimedia Commons) Acetylcholín uvoľňovaný v neuromuskulárnych zväzoch cholinergných receptorov aktívneho kostrového svalu nazývaný nikotínky, pre jeho citlivosť na nikotínový alkaloid a ktoré sa tiež nachádzajú v synapsách ganglionov autonómneho nervového systému (SNA).
Postgangliové neuróny parasympatického rozdelenia tohto systému vyvíjajú svoje funkcie uvoľňovaním acetylcholínu, ktorý pôsobí na muskarínové cholinergné receptory umiestnené v membránoch efektorových buniek a vyvolávajú v nich elektrické modifikácie zmenami permeability v ich iónových kanáloch.
 Chemická štruktúra acetylcholínu neurotransmitera (zdroj: Neurotokeker [verejná doména] prostredníctvom Wikimedia Commons) [TOC]
Chemická štruktúra acetylcholínu neurotransmitera (zdroj: Neurotokeker [verejná doména] prostredníctvom Wikimedia Commons) [TOC]
Štruktúra
Muskarínové receptory patria do rodiny metabotropných receptorov, čo je termín, s ktorým sú určené tie receptory, ktoré nie sú správne iónové kanály, ale proteínové štruktúry, ktoré pri aktivovaní spúšťajú intracelulárne metabolické procesy, ktoré modifikujú aktivitu skutočných kanálov.
Tento termín sa používa na ich odlíšenie od ionotropných receptorov, ktoré sú skutočnými iónovými kanálmi, ktoré sa otvárajú alebo blízko priamym pôsobením neurotransmitera, ako je to v prípade nikotínových receptorov, ktoré sú už pomenované z neuromuskulárnych plakov kostrových svalov.
V rámci metabotropných receptorov sú muskalíny zahrnuté v skupine známe ako štrbina G, aktivujú fosfolipázu C (PLC).
Muskarínové receptory sú dlhé komplexné membránové proteíny; Majú sedem transmarketových segmentov zložených z alfa préfov, ktoré postupne prechádzajú cez lipidovú membránovú dvojvrstvu. Vo vnútri, na cytoplazmatickej tvári, sú spojené s zodpovedajúcim G proteínom, ktorý prenáša interakciu ligand-recepvery.
Môže vám slúžiť: Embrioblasts: vývoj, funkcie a vrstvyTypy muskarínových receptorov a ich funkcie
Bolo identifikovaných najmenej 5 typov muskarínových receptorov, ktoré sú označené písmenom M, nasledované číslo, a to: M1, M2, M3, M4 a M5.
Receptory M1, M3 a M5 tvoria rodinu M1 a vyznačujú sa ich asociáciou proteínov GQ alebo G11, zatiaľ čo receptory M2 a M4 sú z rodiny M2 a sú spojené s proteínom GI proteín.
- M1 receptory
Nachádzajú sa hlavne v centrálnom nervovom systéme, v exokrinných žľazach a v autonómnom gangliách Nodon. Sú spojené s GQ proteínom, ktorý aktivuje enzým fosfolipázy C, ktorý sa premieňa na fosfatidyl -inozitol (PIP2) na inozitol tryposfát (IP3), ktorý uvoľňuje intracelulárny a diascilglycerol (DAG), ktorý aktivuje proteíny C.
- M2 receptory
V zásade sa nachádzajú v srdci, hlavne v bunkách sinoaurikulárneho uzla, na ktorom pôsobia znížením svojej frekvencie výtoku, ako je opísané nižšie.
Srdcový automatizmus
M2 receptory boli dôkladnejšie študované v Nodoauricular Conda (SA) srdca, kde sa normálne prejavuje automatizmus, ktorý pravidelne vytvára rytmické excitácie zodpovedné za srdcovú mechanickú aktivitu.
Bunky sinoaurikulárnej uzliny po každom akčnom potenciáli (PA), ktoré vyvolávajú srdcový systol (kontrakcia), sa repolarizujú a vrátia sa na úroveň asi -70 mV. Napätie však nezostane v tejto hodnote, ale trpí progresívnou depolarizáciou na prahovú úroveň, ktorá spúšťa nový akčný potenciál.
Táto progresívna depolarizácia je spôsobená spontánnymi zmenami iónových prúdov (I), ktoré zahŕňajú: k+ (IK1) redukciu výstupu, vzhľad vstupného prúdu Na+ (FI) a potom vstup Ca ++ (ICAT), až kým nedosiahne prah a spustí sa ďalší prúd Ca ++ (ICal) zodpovedný za akčný potenciál.
Ak je výstup K+ (IK1) veľmi nízky a vstupné prúdy Na+ (IF) a Ca ++ (ICAT) sú vysoké, depolarizácia sa vyskytuje rýchlejšie, akčný potenciál a kontrakcia sa vyskytuje skôr a srdcový frekvencia je vyšší. Opačné úpravy v týchto prúdoch nižšia frekvencia.
Môže vám slúžiť: kariocinesisMetabotropické zmeny vyvolané norepinefrínom (sympatickým) a acetylcholínom (parasympatický) môžu tieto prúdy zmeniť. AMPC priamo aktivuje kanály IF, proteíny A (PKA) fosforyila a aktivuje kanály ICAT Ca ++ a skupina proteínov β gi aktivuje výstup K+.
Muškarinická akcia M2
Keď sa acetylcholín uvoľňoval postgangliovým koncom koncovky srdcových vagálnych vlákien (parasympatické) viaže na m2 muskarínové receptory buniek sinoaurikulárnych uzlín, ai podjednotka proteínu GI mení svoj GDP pomocou GTP a odchádza zadarmo do bloku I βyb βγ.
Podjednotka ai inhibuje adenylciklázu a znižuje produkciu AMPC, čo znižuje aktivitu kanálov pre IF a aktivitu PKA. Táto posledná skutočnosť znižuje fosforyláciu a aktivitu kanálov Ca ++ pre ICAT; Výsledkom je zníženie depolarizačných prúdov.
Skupina vytvorená py podjednotkami GI proteínu aktivuje prúd K+ out (IKACH), ktorý má tendenciu pôsobiť proti vstupom Na+ a Ca ++ a znižuje rýchlosť depolarizácie.
Kĺbovým výsledkom je zníženie sklonu spontánnej depolarizácie a zníženie srdcovej frekvencie.
- M3 receptory
 Schéma m3 muskarínového receptora (zdroj: Takuma-SA [CC0] cez Wikimedia Commons)
Schéma m3 muskarínového receptora (zdroj: Takuma-SA [CC0] cez Wikimedia Commons) Nachádzajú sa v hladkom svale (tráviaci systém, močový mechúr, krvné cievy, bronchi), v niektorých exokrinných žľazach a v centrálnom nervovom systéme.
GQ proteín je tiež spojený a na pľúcnej úrovni môžu spôsobiť bronchokonstrikciu, zatiaľ čo pôsobia vo vaskulárnom oxidu dusnatého uvoľňovania endotelu (NO) a spôsobujú vazodilatáciu.
- Receptory M4 a M5
Tieto receptory sú menej charakterizované a študované ako tie predchádzajúce. Jeho prítomnosť v centrálnom nervovom systéme a v niektorých periférnych tkanivách bola hlásená, ale ich funkcie nie sú jasne stanovené.
Môže vám slúžiť: Tata Box: Charakteristiky a funkcieAntagonista
Univerzálny antagonista týchto receptorov je atropín, alkaloid extrahovaný z rastliny Atropa, To sa na ne viaže s vysokou afinitou, ktorá predstavuje kritérium na ich odlíšenie od nikotínových receptorov, ktoré sú necitlivé na túto molekulu.
Existuje veľké množstvo ďalších antagonistických látok, ktoré sa viažu na rôzne typy muskarínových receptorov s rôznymi afinitami. Kombinácia rôznych hodnôt afinity pre niektoré z nich slúžila presne na zahrnutie týchto receptorov do jednej alebo druhej z opísaných kategórií.
Čiastočný zoznam ďalších antagonistov by zahŕňal: pirenzepín, metolektrémín, 4-DAMP, hymbacín, AF-DX 384, Tripitramín, Darifenacín, PD 102807, AQ RA 741, PFHHSID, MT3 a MT7; Toxíny, ktoré boli uvedené v jedoch zelených a čiernych mambákov, respektíve.
Napríklad receptory M1 majú vysokú citlivosť na pirenzepín; M2 Tripitramínom, metctramínom a hybakínom; M3 po 4-DAMP; M4 veľmi súvisí s toxínom MT3 a tiež s hybakínom; M5 sú veľmi podobné M3, ale s ohľadom na nich sú menej príbuzní tu 741.
Odkazy
- Ganong WF: Neurotransmitery a neuromodulátory, In: Preskúmanie lekárskej fyziológie, 25. vydanie. New York, McGraw-Hill Education, 2016.
- González JC: Úloha muskarínových receptorov pri modulácii GABAergického prenosu v hippocampe. Pamätajte, že sa rozhodnete pre doktora. Autonómna univerzita v Madride. 2013.
- Guyton AC, Hall JE: Rythmické vzrušenie srdca, In: Učebnica lekárskej fyziológie , 13. vydanie; AC Guyton, JE Hall (eds). Philadelphia, Elsevier Inc., 2016.
- Piper HM: Herzerregung, In: Physiologie des Menschen Mite patofysiologie, 31. vydanie; RF Schmidt a kol. (Eds). Heidelberg, Springer Medizin Verlag, 2010.
- Schrader J, Gödeche A, Kelm M: Das Hertz, In: Fyziológia, 6. vydanie; R Klinke a kol. (Eds). Stuttgart, Georg Thieme Verlag, 2010.
- Siegelbaum SA, Clafam DE, Schwartz JH: Modulácia synaptického prenosu: Druhá poslovia, In: Princípy nervovej vedy, 5. vydanie; E Kandel a kol. (Eds). New York, McGraw-Hill, 2013.
- « História bunkovej biológie, aké štúdie, aplikácie a koncepty
- Ako sa rastliny reprodukujú? Asexuálna a sexuálna reprodukcia »