

Charakteristiky mikrofilantov, štruktúra, funkcie, patológia
- 4954
- 200
- Adrián Fajnor
Ten mikrofilamenty o Actínové vlákna, sú jednou z troch základných zložiek eukaryotických buniek cytoskelet.
V eukaryotoch sú gény kódujúce aktínové mikrofilamenty vysoko zachované vo všetkých organizmoch, takže sa často používajú ako molekulárne markery pre rôzne štúdie.
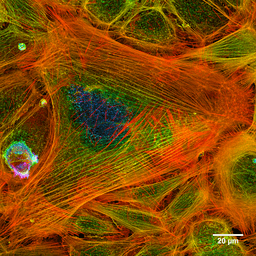
Fotografia aktínových vlákien zafarbenej bunky (Zdroj: Howard Vindin [CC BY-SA 4.0 (https: // creativeCommons.Org/licencie/By-SA/4.0)] Via Wikimedia Commons) Mikrofilamenty sú distribuované v celom cytosóle, ale sú obzvlášť hojné v podkladovej oblasti do plazmatickej membrány, kde tvoria komplexnú sieť a sú spojené s inými špeciálnymi proteínmi za vzniku cytoskeletu.
Mikrodilamentové siete v cytoplazme cicavčích buniek sú kódované dvoma zo šiestich génov opísaných pre aktín, ktoré sú zapojené do dynamiky mikrofilamentov a ktoré sú dokonca veľmi dôležité počas diferenciácie kmeňových buniek.
Mnoho autorov súhlasí s tým, že mikrofilamenty sú najrôznejšie, univerzálne a najdôležitejšie proteíny cytoskeletu väčšiny eukaryotických buniek a je dôležité si uvedomiť, že sa nenachádzajú v prokaryotických mikroorganizmoch.
V tomto type buniek však existujú homológne vlákna s mikrofilanciami, ale ktoré sú tvorené iným proteínom: proteín MREB.
V súčasnosti sa predpokladá, že gén, ktorý kóduje tento proteín, je možným génom predkov pre eukaryotický aktín. Avšak homológia aminokyselinovej sekvencie, ktorá tvorí proteín MREB, je iba 15% vzhľadom na aktínovú sekvenciu.
Rovnako ako základná súčasť cytoskeletu, akýkoľvek fenotypový defekt v mikrotubuloch, ako aj v stredných vláknach a aktínových mikrofilamentoch (cytoskelet) môže spôsobiť rôzne bunkové a systémové patológie.
[TOC]
Vlastnosti
Mikrofilamenty sa skladajú z proteínových monomérov rodiny aktínov, ktoré sú mimoriadne hojné kontraktilné proteíny v eukaryotických bunkách, pretože sa tiež podieľajú na kontrakcii svalov.
Tieto vlákna majú priemer medzi 5 a 7 nm, takže sú známe aj ako tenké vlákna a sú zložené z dvoch foriem aktínu: globulárna forma (aktín g) a vláknitý tvar (aktín f).
Proteíny, ktoré sa zúčastňujú na cytoskelete, sú známe ako y a β aktíny, zatiaľ čo protesty, ktoré sa zúčastňujú na kontrakcii, sú zvyčajne aktíny a.
Môže vám slúžiť: cytoplazma: funkcie, časti a vlastnostiPodiel globulárneho aktínu a vláknitého aktínu v cytosóle závisí od bunkových potrieb, pretože mikrofilancie sú veľmi variabilné a všestranné štruktúry, ktoré neustále rastú a skracujú v dôsledku polymerizácie a depolymetrizácie.
Actín g je malý guľový proteín, ktorý sa skladá z takmer 400 aminokyselín a približne 43 kDa molekulovej hmotnosti.
Actín g monoméry, ktoré tvoria mikrofilamenty, sú usporiadané vo forme špirálového vlákna, pretože každý z nich trpí objatím, keď je spojený s nasledujúcim.
Actín G je spojený s molekulou Ca2+ a ďalším ATP, ktorý stabilizuje jeho guľový tvar; Zatiaľ čo aktín F sa získa po hydrolýze terminálneho fosfátu molekuly ATP v aktíne, čo prispieva k polymerizácii.
Organizácia
Actínové vlákna môžu byť usporiadané vo forme „lúčov“ alebo „sietí“, ktoré majú vo vnútri buniek rôzne funkcie. Lúče tvoria paralelné štruktúry spojené pomerne tuhými krížovými mostmi.
Siete, na druhej strane, sú pohodlnejšie štruktúry, napríklad tri rozmerové oká s vlastnosťami polo -pevných gélov.
Existuje mnoho proteínov, ktoré sú spojené s aktínovými alebo mikrofilamentovými vláknami a ktoré sú známe ako ABP (z angličtiny Proteíny viažuce sa na aktín), ktoré majú na to konkrétne stránky.
Mnohé z týchto proteínov umožňujú mikrofilancie interagovať s ďalšími dvoma zložkami cytoskeletu: mikrotubuly a stredné vlákna, ako aj s ostatnými zložkami vnútornej tváre plazmatickej membrány.
Medzi ďalšie proteíny interagujú s mikrofilanciami patria jadrové listy a spektín (v červených krvinkách).
Ako sa tvoria aktínové vlákna?
Pretože guľové aktínové monoméry sa vždy spájajú rovnakým spôsobom, orientované rovnakým smerom, mikrofilamenty majú definovanú polaritu s dvoma extrémami: jeden „viac“ a jeden „menej“.
Polarita týchto vlákien je veľmi dôležitá, pretože z dôvodu ich pozitívneho konca rastú podstatne rýchlejšie, kde sa pridávajú nové aktívne monoméry.
 Grafické znázornenie aktínových mikrofilantov (Zdroj: Derivátová práca: Retama (talk) Thin_Filament_formation.SVG: Mikael Häggström [verejná doména] cez Wikimedia Commons)
Grafické znázornenie aktínových mikrofilantov (Zdroj: Derivátová práca: Retama (talk) Thin_Filament_formation.SVG: Mikael Häggström [verejná doména] cez Wikimedia Commons) Prvá vec, ktorá sa odohráva počas polymerizácie aktínových vlákien, je proces známy ako „nukleácia“, ktorý pozostáva z asociácie troch proteínových monomérov.
Môže vám slúžiť: plazomolýzaK tomuto zastrihávači sa nové monoméry pridávajú oboma koncami, takže vlákno rastie. Actínové monoméry sú schopné hydrolyzovať ATP s každým zväzkom, ktorý má dôsledky na rýchlosť polymerizácie, pretože časti aktínu-ATP disociujú väčšie ťažkosti ako aktín-adp.
ATP nie je potrebný na polymerizáciu a betónová funkcia jeho hydrolýzy ešte nebola objasnená.
Niektorí autori sa domnievajú, že keďže aktínové polymerizačné udalosti sú rýchle.
Regulácia
Polymerizácia aktínových vlákien a jeho depolimerizácia sú procesy vysoko regulované radom špecifických proteínov, ktoré sú zodpovedné za prestavbu vlákien.
Príklad proteínov, ktoré regulujú depolyméry. Ďalší proteín, profylaín, má opačnú funkciu, pretože stimuluje spojenie monomérov (stimuláciou výmeny ADP za ATP).
Funkcia
Mikrofilamenty interagujú s myozínovými vláknami, ktoré sú spojené s transmembránovými proteínmi, ktoré majú doménu v cytosóle a ďalšej zahraničnej bunke, takže sa zúčastňujú procesov mobility buniek.
Tieto mikrofilamenty spojené s plazmatickou membránou sprostredkujú rôzne bunkové reakcie na rôzne druhy stimulov. Napríklad bunková adhézia epitelových tkanív je riadená transmembránovými proteínmi známymi ako kadheríny, ktoré interagujú s mikrofilanciami na nábor faktorov odpovedí.
Actínové vlákna interagujú so strednými vláknami, aby sa extracelulárne stimuly vysielali na kľúčové miesta, ako sú ribozómy a chromozómy vo vnútri jadra.
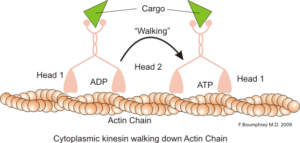 Reprezentácia intracelulárnej motorickej funkcie aktínových mikrofilancií (zdroj: boumpreyfr [CC BY-SA 3.0 (https: // creativeCommons.Org/licencie/By-SA/3.0)] Via Wikimedia Commons)
Reprezentácia intracelulárnej motorickej funkcie aktínových mikrofilancií (zdroj: boumpreyfr [CC BY-SA 3.0 (https: // creativeCommons.Org/licencie/By-SA/3.0)] Via Wikimedia Commons) Klasická a veľmi študovaná funkcia mikrofilantov je jej schopnosť vytvárať „mosty“, „koľajnice“ alebo „diaľnice“ pre pohyb myozínového motorického proteínu I, ktorý je schopný načítať transportné vezikuly z organelov do membránovej plazmy na sekrečnom sektore dráha.
Mikrofilamenty tiež interagujú s myozínom II na vytvorenie kontraktilného kruhu, ktorý sa tvorí počas cytokinézy, presne počas poslednej fázy bunkového delenia, v ktorom je cytosol oddelený od kmeňových a dcérskych buniek.
Môže vám slúžiť: polymorfonukleárne leukocytyVšeobecne platí, že aktínové mikrofilamenty modulujú distribúciu niektorých organel, ako je Golgiho komplex, endoplazmatické retikula a mitochondrie. Okrem toho sa tiež podieľajú na priestorovom umiestnení RNM, takže ich čítajú ribozómy.
Celá sada mikrofilantov mobilných telefónov, najmä tie, ktoré úzko súvisia s plazmatickou membránou, sa podieľajú na tvorbe zvlnených bunkových membrán buniek, ktoré majú konštantný aktívny pohyb.
Zúčastňujú sa tiež na tvorbe mikrovingov a iných spoločných výbežkov na povrchu mnohých buniek.
Príklad funkcií v pečeni
Mikrofilamenty sa podieľajú na procese vylučovania žlčových škôd v hepatocytoch (pečeňové bunky) a tiež na peristaltických pohyboch (koordinovaná kontrakcia) pečeňových kanálikov.
Prispievajú k diferenciácii domén plazmy membrány vďaka ich spojení s rôznymi cytosolickými prvkami a kontrole, ktorú vyvíjajú na topografiu týchto intracelulárnych prvkov.
Súvisiace patológie
S primárnym defektom v štruktúre alebo s proteínmi a regulačnými enzýmami v syntéze mikrofilantov je spojených málo chorôb, hoci sú priamo zapojené do veľkého počtu funkcií.
Nízky index chorôb a malformácií v primárnej štruktúre mikrofilancií je spôsobený skutočnosťou, že vo všeobecnosti existujú viacnásobné kódujúce gény aktínu a jeho regulačných proteínov, čo je jav známy ako „genetická redundancia“.
Jednou z najviac študovaných patológií je vitrifikácia oocytov na ich cytoskelete, kde sa pozoruje prerušenie v sieti kortikálnych mikrofilamentov, ako aj depolimerizácia a dezorganizácia mikrotubulov mitotického vretena.
Všeobecne povedané, táto vitrifikácia spôsobuje chromozomálnu disperziu, pretože vedie k nezhode v zhutnení celého chromatínu.
Bunky, ktoré majú väčšiu organizáciu a podiel mikrofilantov vo svojom cytoskelete, sú pruhované svalové bunky, preto je väčšina patológií spojená s poruchou kontraktilného prístroja.
Defektné alebo atypické mikrofilamenty boli tiež spojené s ochorením kostí známe ako ochorenie paget.
Odkazy
- Aguilar-Cuenca, R., Llorente-González, C., Vicente, C., & Vicente-Manzanares, m. (2017). Dynamika adhézie koordinovanej mikrofilamentom vedie k migrácii jednotlivých buniek a tvarmi, ktoré tkanivá. F1000Resarch, 6.
- Dve prostriedky, c. G., Chhabra, D., Kekic, m., Prst, ja. Vložka., Tsubakihara, m., Berry, D. Do., & Nosworthy, n. J. (2003). Väzbové proteíny aktín: regulácia cytoskeletálnych mikrofilancií. Fyziologické recenzie, 83(2), 433-473.
- Guo, h., Fauci, l., Shelley, m., & Kanso a. (2018). Bistabilita v synchronizácii ovládacích mikrofilantov. Journal of Fluid Mechanics, 836, 304-323.
- Spustiť., Langer, r., & Vacanti, J. P. (Eds.). (2011). Princípy tkanivového inžinierstva. Akademická tlač.
- Robbins, J. (2017). Choroby cytoskeletu: Deminopatie. V kardioskeletálnych myopatiách u detí a dospelých (PP. 173-192). Akademická tlač.
- « Charakteristiky grizzly medveďov, biotop, jedlo, správanie
- Cistus clusii Charakteristiky, taxonómia, biotop, distribúcia, použitia »